Morphologie

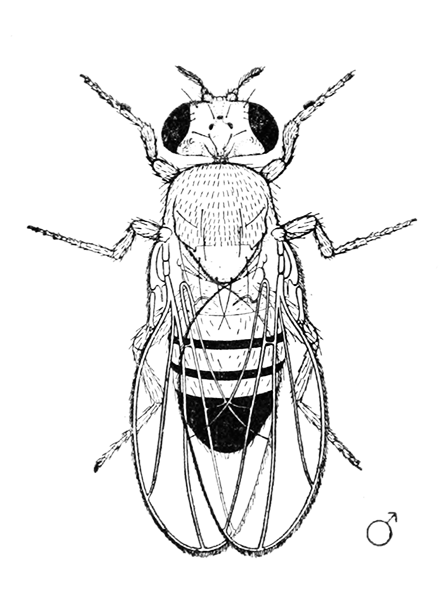
Drosophila melanogaster, images tirées de : Morgan T. H.. 1919. The Physical Basis of heredity. Philadelphia, J.B. Lippincott Company.
Le male a un peigne sexuel sur la première paire de pattes, est plus petit que la femelle, l'extémité de son abdomen est plus sombre et arondie.
Phénotypes et génotypes

[corps noir]. La mutation du gène black (b) renforce la couleur noire du corps par accumulation de mélanine; l'allèle muté est récessif.

[yeux réduits]. La mutation du gène bar (B) réduit le nombre de facettes (ommatidies): il chute de 750 facettes à 80 chez les individus mutés homozygotes. Les femelles hétérozygotes possèdent environ 350 facettes. Le gène est lié au sexe.

[yeux bruns]. La mutation du gène brown (bw) fait disparaitre le pigment rouge normalement présent dans l'œil qui devient rouge brun; ce gène entre en interaction avec d'autres gènes contrôlant la couleur de l'œil. L'allèle muté est récessif.

[yeux rouge vif]. La mutation du gène cinnabar (cn) fait disparaitre le pigment brun normalement présent dans l'œil qui devient rouge clair; ce gène entre en interaction avec d'autres gènes contrôlant la couleur de l'œil. L'allèle muté est récessif.
[ailes frisées]. La mutation du gène curly (Cy) recourbe les ailes ce qui empêche les mouches de voler; l'allèle muté est dominant (les ailes sont moyennement frisées chez les hétérozygotes). Les mouches homozygotes pour cet allèle sont naines et fertiles mais leur taux de survie est très faible (on dit que cette mutation est létale ou quasi létale).
[corps ébène]. La mutation du gène ebony (e) renforce la couleur noire (ébène) du corps par accumulation de mélanine; l'aspect est très proche de celui provoqué par la mutation du gène b, mais le comportement sexuel est davantage perturbé; l'allèle muté est récessif.
[yeux rouge vif]. La mutation du gène scarlet (st) fait disparaitre le pigment brun normalement présent dans l'œil qui devient rouge vif; ce gène entre en interaction avec d'autres gènes contrôlant la couleur de l'œil. L'allèle muté est récessif.

[ailes vestigiales]. La mutation du gène vestigial (vg) réduit l'aile à un moignon; l'allèle muté est récessif.

[yeux blancs]. La mutation du gène white (w) fait perdre les pigments de l'oeil qui devient blanc; ce gène entre en interaction avec d'autres gènes contrôlant la couleur de l'œil, les mouches homozygotes pour l'allèle muté ont toujours les yeux blancs quelque soit l'état des autres gènes contrôlant la couleur de l'œil. L'allèle muté est récessif et le gène lié au sexe.

[corps jaune]. La mutation du gène yellow (y) empêche la mouche de fabriquer le pigment noir de son corps; le comportement sexuel du mâle est perturbé; l'allèle muté est récessif est le gène est lié au sexe. Dans le simulateur, l'interaction des gènes yellow avec les gènes black ou ebony a été simplifiée.
Les images de Drosophile illustrant les phénotypes mutés ci-dessus sont à créditer à l'Exploratorium de San Francisco (Etats-Unis). Elles ont été reprises avec plus ou moins de modifications dans les deux premières versions de l'application Droso et possèdent leurs propres droits d'usage.
↑ Exploratorium.
Site du musée scientifique de San Francisco (USA); la source première des images utilisées par Droso 1 et 2.
Site du musée scientifique de San Francisco (USA); la source première des images utilisées par Droso 1 et 2.
Gènes liés et carte génétique
tous les gènes ne sont pas indépendants
Tous les caractères ne sont pas indépendants comme l'avait énoncé Gregor Mendel, mais certains ont tendance à rester ensemble dans la descendance. Les études de dihybridisme permettent de préciser les liaisons entre caractère (gènes). Dans l'étude de deux gènes (dihybridisme), lorsque les 4 types de gamètes produits par les hybides F1 sont équiprobables, les gènes sont indépendants. Dans le cas contraire (les combinaisons d'allèles qui n'existaient pas chez les parents sont chacune inférieure à 25%), les gènes sont liés. Ces relations entre gènes liés permettent de constituer des groupes de liaison.les chromosomes jouent un rôle déterminant dans l'hérédité
On s'est aperçu ensuite que le nombre de groupes de liaison correspondait au nombre de chromosomes, ce qui a constitué un argument (mais pas le seul) pour établir le modèle chromosomique de l'hérédité. Chez la Drosophile il existe quatre chromosomes (plus précisément quatre paires de chromosomes homologues) et quatre groupes de liaison. Les gènes liés sont présents sur le même chromosome et leur organisation est linéaire (ce qui s'explique par la structure de l'ADN).
Groupes de liaisons (et chromosomes) de la Drosophile
Les nombres représentent les distances génétiques en cM.
Le chromosome Y (non représenté) est pratiquement dépourvu de gènes.
Pour établir une carte génétique, on établit les distances entre gènes proches, puis on additionne les distances. L'unité de mesure est le centiMorgan (cM), en hommage à Thomas Morgan l'un des généticiens ayant mis en évidence les groupes de liaisons.
Les gènes d'un même chromosome étant physiquement liés, on ne devrait pas avoir de recombinés du tout entre gènes liés. La présence des recombinés s'explique par le mécanisme nommé crossing-over (plus ou moins bien traduit par enjambement). Les deux chromosmes homologues (ceux qui portent les mêmes gènes mais pas forcément les mêmes allèles se regroupent par paire lors de la méiose et peuvent échanger des segments. Ces échanges permutent les allèles. Les gènes étant disposés linéairement sur le chromosome, plus leur distance est grande et plus les échanges peuvent se produire. Mais on ne peut dépasser le pourcentage d'échanges (on dit aussi brassages) obtenus avec des gènes disposés sur des chromosomes différents qui est de 50%. C'est pourquoi la proportion de recombinés ne croit pas proportionnellement à la distance génétique mais se rapproche d'une asymptote à 50%. Une façon alternative d'expliquer ce fait est de constater que des crossing-over multiples peuvent se produire lorsque la distance est grande; dans ce cas seul un nombre impair de crossing-over permute les allèles donc avec une grande distance il est impossible d'avoir plus de 50% de recombinaisons (le nombre pair de crossing-over pair est alors proche du nombre impair de crossing-over).
Dans la pratique lorsque deux gènes ont une fréquence de recomninaison inférieure à 50%, ils sont dans le même groupe de liaison. Si la fréquence est égale à 50% on recherche un gène intermédiaire. S'il n'y a pas de gène intermédiaire, les gènes ont de fortes chances de se trouver dans des groupes de liaisons différents.
chaque groupe de liaison correspond à une paire de chromosomes homologues
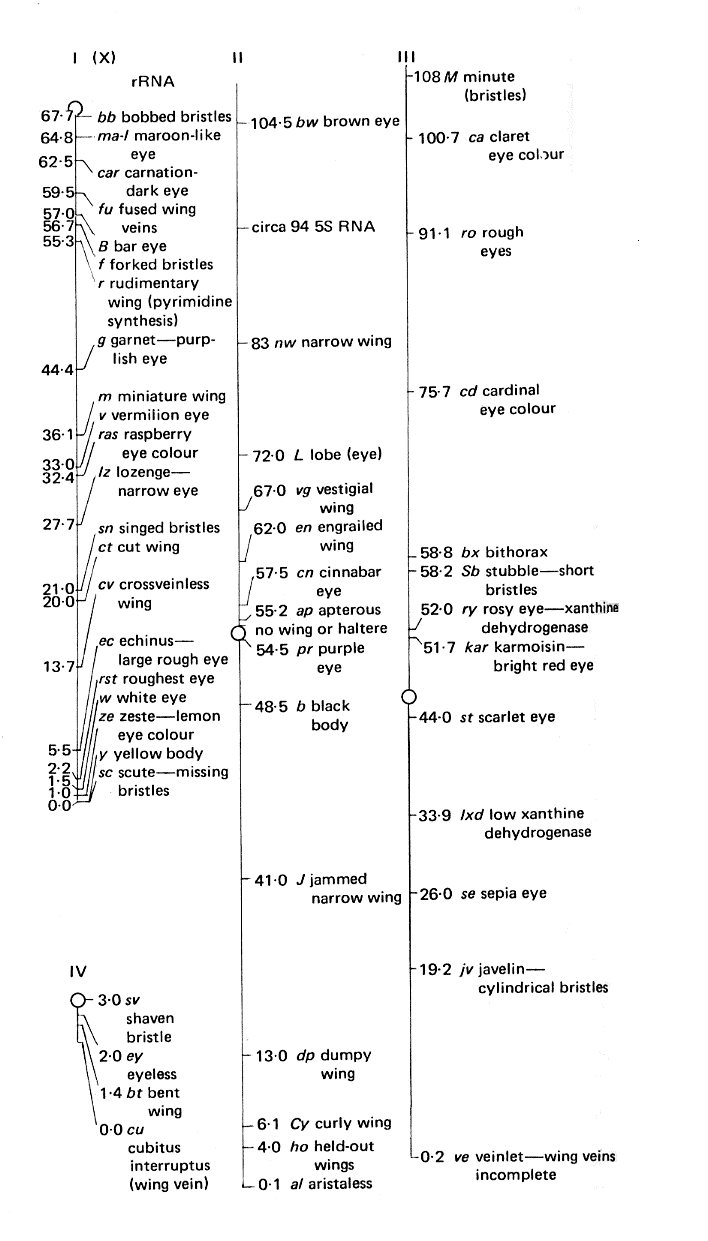
Les valeurs sont données en cM et mesurées à partir du gène le plus proche d'une extrémité. Des grandes valeurs sont calculées comme des sommes d'intervalles courts car la fréquence de recombinaison pour deux loci ne peut pas dépasser 50%.
Références
Adresse de cette page: http://www.didac-tic.fr/droso/docs/index.php