- Trois exemples iconiques
- Résistances
- La domestication des plantes
- La domestication du Maïs (page développée)
- La domestication du Riz
- Plantes OGM
Il est possible d'observer l'évolution sur des temps relativement courts, à l'échelle d'une décennie ou de quelques années. Pour des êtres vivants dont le temps de reproduction est très court comme les bactéries, cela peut même se compter en semaines. Leur rapidité (relative) peut s'expliquer par la constance de la sélection orientée dans une direction toujours la même pendant une période de temps suffisant. Au contraire dans la nature la sélection est souvent fluctuante, ne générant qu'une sorte de bruit de fond évolutif que Stephen Jay Gould, dans le cadre du modèle des équilibres ponctués qualifierait de stagnation. Il est possible d'observer des sélections sous l'effet des fluctuations climatiques naturelles, comme chez des Pinsons des Galàpagos. L'exemple de la Phalène, est causée par une modification humaine de l'environnement, mais c'est une sélection ponctuelle et involontaire, une sorte de dommage collatéral.
D'autres transformations crées de la main de l'Homme nous impliquent bien davantage. Mais l'état final n'est pas toujours celui souhaité; il peut interroger notre prétention à changer le monde et annuler ce que nous avions considéré, dans un premier temps, comme des avancées extraordinaires (nous affranchir d'insectes ravageurs de nos cultures, d'insectes vecteurs de maladies, de redoutables infections bactériennes, produire des ressources alimentaires indépendantes du reste du vivant, etc.). L'échec était pourtant prévisible.
Trois exemples iconiques ↑
La Phalène (Biston betularia), (Peppered Moth) ↑
Un exemple emblématique: le mélanisme industriel
Dans les régions industrielles anglaises la variété sombre du papillon était devenue dominante dès la fin du 19e siècle, alors que personne ne l'avait remarquée au début du même siècle. Dans les mêmes régions qui se sont désindustrialisées et assainies depuis, la fréquence a diminué pour devenir quasi nulle à la fin du 20e siècle.
Biston betularia typica (Lodz, Pologne); Crédit: Jerzy Strzelecki, Wikimedia, GNU free documentation licence, CC by SA.
- il est spectaculaire;
- une explication intuitive est communément partagée;
- notre intuition pourrait être fausse, mais les preuves à l'appui de cette explication sont nombreuses.
L'exemple a pourtant sucité des débats passionnés chez les scientifiques (sans compter les créationnistes qui ont essayé d'exploiter ces désaccords) que certains ont résumé, non sans jeu de mots, par: «La Phalène du bouleau, une histoire en noir et blanc» (Jim Mallet, 2003).

Glasgow, vers 1880; gravure sur bois, auteur inconnu, domaine public.

Biston betularia carbonaria (Lodz, Pologne); Crédit: Jerzy Strzelecki, Wikimedia, GNU free documentation licence, CC by SA.
L'espace et le temps
On avait constaté vers le mileu du 20e siècle que la fréquence de la variété sombre était beaucoup plus élevée dans les régions polluées (troncs d'arbres couverts de suie et dépourvus de lichens) que dans celles qui ne l'étaient pas. Ce que suggère notre représentation du monde à l'observation des photographies se produit-il réelllement dans la nature ? autrement dit, l'évolution constatée des fréquences des deux formes les plus nombreuses du papillon (carbonaria et typica) s'explique-t-elle par la pression de sélection exercée par une prédation différentielle des oiseaux ? 
Biston betularia insularia (Warwickshire, England); © Paul Nicholas.
Un aspect totalement passé sous silence dans les citations de l'expérience cruciale de Kettlewell (mais pas par son auteur !) est l'existence de formes intermédaires entre typica et carbonaria. Nommées insularia, elles sont peu nombreuses dans la région de Birmingham lors de l'expérience (1). Contrairement à ce qu'on pourrait croire spontanément, ces phénotypes n'ont rien à voir avec une dominance partielle du caractère carbonaria. Le fait est pris en compte par Kettlewell qui incorpore ces formes intermédiaires dans son protocole. Ces formes présentent une grande variabilité, et il n'y a guère que le corps qui reste totalement noir. Il s'agit en fait d'un ensemble hétérogène (cf plus bas avec l'oeil des généticiens).
Une des premières expériences consistait à lâcher des Phalènes des deux variétés dans une grande cage contenant des troncs d'arbres clairs et foncés puis d'y introduire des Mésanges (un protocole un peu biaisé), mais plus tard Kettlewell réalise des expériences en milieu naturel: des Phalènes mâles (2) des trois variétés, marquées par une petite tâche de couleur en dessous du corps sont lâchés dans un bois proche de Birmingham et pollué par la suie.
nombre de papillons
variété |
lâchés | recapturés |
|---|---|---|
| carbonaria | 447 | 123 (21%) |
| typica | 137 | 18 (11%) |
| insularia | 46 | 8 (15%) |
Une contre-épreuve organisée en zone non polluée à montré que cette fois ce sont les papillons carbonaria qui sont le plus capturés par les oiseaux.
Certains détails de l'expérience ont été critiqués (sous l'influence de créationnistes qui supportaient mal cet exemple emblématique): par exemple, le grand nombre de papillons relâchés peut modifier le comportement de prédation des oiseaux. Et sur des temps plus longs, les migrations entre régions polluées et non polluées peuvent diminuer l'effet de la prédation. Laurence Cook (Cook, 2013) tout en concluant que la prédation semble jouer le rôle principal cite quelques autres explications possibles de l'évolution dont au moins une était évoquée par Kettlewell lui-même dans d'autres articles: des vigueurs différentes présentées par chacune des deux variétés les plus nombreuses et des hybrides de ces deux variétés.
(1)
Le caractère carbonaria dominant tous les autres dans les hybridations, sa forte fréquence avait tendance à masquer les allèles et gènes déterminant le caractère insularia chez les hybrides. La baisse de la fréquence de carbonaria donne aujourd'hui nettement plus d'importance à insularia qui peut représenter, dans certaines zones 30% des phénotypes des Phalènes.
(2) Il est très difficile de recapturer des femelles en les attirant avec des lampes c'est pourquoi Kettlewell ne lâche que des mâles.
↑ Peppered moth evolution. Wikipedia (en).
La page n'existe pas en français.
↑ Henry Bernard Kettlewell. 1955. Selection experiments on industrial melanism in the Lepidoptera. Heredity 9: 323–342. DOI: 10.1038/hdy.1955.36
↑ Henry Bernard Kettlewell. 1956. Further selection experiments on industrial melanism in the Lepidoptera. Heredity 10 (3): 287-301.
↑ Bruce S. Grant. 1999. Fine tuning the peppered Moth paradigm. Evolution, 53: 980-984. DOI: 10.1111/j.1558-5646.1999.tb05394.x
(2) Il est très difficile de recapturer des femelles en les attirant avec des lampes c'est pourquoi Kettlewell ne lâche que des mâles.
↑ Peppered moth evolution. Wikipedia (en).
La page n'existe pas en français.
↑ Henry Bernard Kettlewell. 1955. Selection experiments on industrial melanism in the Lepidoptera. Heredity 9: 323–342. DOI: 10.1038/hdy.1955.36
↑ Henry Bernard Kettlewell. 1956. Further selection experiments on industrial melanism in the Lepidoptera. Heredity 10 (3): 287-301.
↑ Bruce S. Grant. 1999. Fine tuning the peppered Moth paradigm. Evolution, 53: 980-984. DOI: 10.1111/j.1558-5646.1999.tb05394.x
Les Pinsons des Galápagos ↑
Hommage à Charles Darwin
Les Pinsons des Galápagos, popularisés par David Lack sous le nom de Pinsons de Darwin (Lack, 1939), ont été classés en un peu moins d'une vingtaine d'espèces, toutes endémiques de ces iles. Charles Darwin en a collecté quelques exemplaires (en fait semble-t-il surtout son assistant) lors du second passage du Beagle aux Galápagos (le premier pour Darwin); contrairement à la légende, Darwin n'est pas devenu évolutionniste en observant ces Pinsons et il n'avait même pas noté sur quelle ile chaque exemplaire avait été collecté (il le fera pour les Oiseaux moqueurs); mais l'hommage est justifié parce que seul le modèle darwinien a permis d'expliquer (bien plus tard) la diversité des Pinsons.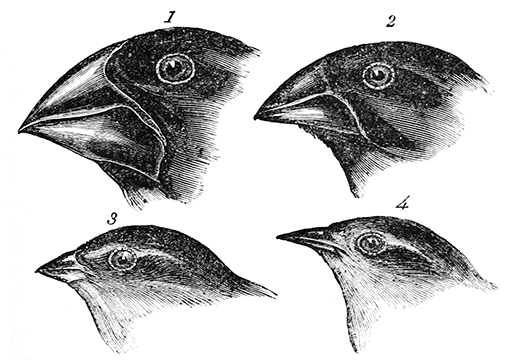
Plate 81: 1. Geospiza magnirostris, 2. Geospiza fortis, 3. Geospiza parvula, 4. Certhidea olivacea; tiré de Darwin, 1845

Héritabilité de la taille* du bec chez G. fortis mesurée en 1976 et en 1978, d'après Grant.
* le terme anglais utilisé est deep et il faudrait sans doute le traduire par "hauteur" plutôt que par "taille".
* le terme anglais utilisé est deep et il faudrait sans doute le traduire par "hauteur" plutôt que par "taille".
Une sélection oscillante
La taille du bec présente toujours une grande variabilité (mise en évidence sur le document par la dispersion des points, et plus forte semble-t-il en 1976), mais en 1978, les becs de petite taille ont disparu chez les parents comme chez la descendance. La pente de la droite (de régression linéaire) est proche de 1, ce qui indique une très forte héritabilité.Les Pinsons G. fortis au grand bec ont mieux survécu (sélection naturelle), de plus les parades précédant l'accouplement (le chant ?) étaient de meilleure qualité et ces mâles étaient davantage choisis par les femelles (sélection sexuelle).
Mais la météorologie changeant constamment d'une année sur l'autre, cette sélection est oscillante (sauf évidemment si le climat change sur le long terme dans une direction déterminée).

Le cratère de Daphne major; Auteur: Erik Enbody © CC by SA.

noir plein: Geospiza fortis, hachré: Geospiza fuliginosa; d'après
Schluter, 1985 (modifié)
Sur Santa Cruz une ile plus grande offrant plus de diversité de nourriture et d'habitat la concurence entre les deux espèces les oblige à se spécialiser davantage (surtout G. fuliginosa sur les petites graines). Sur les iles où les deux espèces sont seules leur source de nourriture est pratiquement la même et la moyenne des tailles de bec se rapproche. Le "déplacement de caractère" avait déjà été noté par David Lack (Lack, 1939, fig.17, p82); mais Lack refusait de lui donner la mondre explication adaptative (Boag & Grant, 1984). Schulter, au contraire de Lack, en s'appuyant sur un traitement mathématique (non repris ici) l'explique par la sélection naturelle (et non par un effet fondateur). Lack changera d'avis dans les rééditions de Darwin's finches.
Sur Daphne, il convient cependant de noter que le Grand Pinson terrestre (G. magnirostris), immigrant occasionnel, est devenu résident à partir de 2005 (Grant,) et que tous les Pinsons peuvent s'hybrider même si les descendances hybrides semblent peu fertiles (Boag, 1981); mais les hybrides G. fortis x G. conirostris semblent faire exception. Un mâle G. conirostris est arrivé en 1981 en provenance d'Española (la seule ile où est présent G. conirostris) et ses descendants hybrides tendent à s'isoler des autres Pinsons de Daphne du point de vue de la reproduction (sans doute à cause du chant et de la différence de taille), constituant potentiellement une nouvelle espèce.
D'une part la classification traditionnelle en différentes espèces des Pinsons de Darwin est sans doute en partie à revoir (Lamichhaney, 2018), d'autre part on ne peut écarter des transferts d'allèles entres espèces (hybridables) et variétés (ces transferts peuvent avoir des conséquences importantes dans la spéciation).
"The remaining land-birds form a most singular group of finches, related to each other in the structure of their beaks, short tails, form of body and plumage: there are thirteen species which Mr. Gould has divided into four sub-groups. All these species are peculiar to this archipelago. (...)
Seeing this gradation and diversity of structure in one small, intimately related group of birds, one might really fancy that from an original paucity of birds in this archipelago, one species had been taken and modified for different ends."
Darwin, 1890 (1845): 378.
Seeing this gradation and diversity of structure in one small, intimately related group of birds, one might really fancy that from an original paucity of birds in this archipelago, one species had been taken and modified for different ends."
Darwin, 1890 (1845): 378.
«Les oiseaux terrestres restants forment un groupe de Pinsons des plus singuliers, apparentés par la structure de leur bec, leur queue courte, la forme de leur corps et leur plumage : on compte treize espèces que M. Gould a divisées en quatre sous-groupes. Toutes ces espèces sont propres à cet archipel. (...)
En constatant cette gradation et cette diversité de structures au sein d'un petit groupe d'oiseaux intimement apparentés, on pourrait croire qu'une espèce, issue d'une rareté originelle d'oiseaux dans cet archipel, a été prélevée et modifiée à des fins différentes.»
En constatant cette gradation et cette diversité de structures au sein d'un petit groupe d'oiseaux intimement apparentés, on pourrait croire qu'une espèce, issue d'une rareté originelle d'oiseaux dans cet archipel, a été prélevée et modifiée à des fins différentes.»
↑ Charles Darwin. 1890 (1845). Journal of Researches into the Natural History and Geology of the Countries visited during the Voyage of H.M.S. Beagle round the World renommé au cours des éditions successives A Naturalist's Voyage Round the World The Voyage Of The Beagle (eleventh edition). John Murray, London.
↑ David Lack. 1939. Darwin's Finches. Harper and Brothers Publishers.
↑ Peter T. Boag. 1981 Morphological variation in the Darwin's Finches (Geospizinae) of Daphne Major Island, Galapagos. Thesis for Peter Grant. DOI: 10.13140/2.1.3688.8329
↑ Peter T. Boag. 1983. The heritability of external morphology in Drawin's Ground Finches (Geospiza) on isla Daphne Major, Galapagos. The Society for the Study of Evolution. DOI: 10.1111/j.1558-5646.1983.tb05618.x
↑ Dolph Schluter et al.. 1985. Ecological Character Displacement in Darwin's Finches. Science 227: 1056-1058. DOI: 10.1126/science.227.4690.1056.
↑ Peter R. Grant and B. Rosemary Grant. 2002. Unpredictable Evolution in a 30-Year Study of Darwin’s Finches. Science vol 296: 707-711.
↑ McNew et al.. 2017. Epigenetic variation between urban and rural populations of Darwin’s finches. BMC Evol Biol 17, 183. DOI: 10.1186/s12862-017-1025-9
↑ David Lack. 1939. Darwin's Finches. Harper and Brothers Publishers.
↑ Peter T. Boag. 1981 Morphological variation in the Darwin's Finches (Geospizinae) of Daphne Major Island, Galapagos. Thesis for Peter Grant. DOI: 10.13140/2.1.3688.8329
↑ Peter T. Boag. 1983. The heritability of external morphology in Drawin's Ground Finches (Geospiza) on isla Daphne Major, Galapagos. The Society for the Study of Evolution. DOI: 10.1111/j.1558-5646.1983.tb05618.x
↑ Dolph Schluter et al.. 1985. Ecological Character Displacement in Darwin's Finches. Science 227: 1056-1058. DOI: 10.1126/science.227.4690.1056.
↑ Peter R. Grant and B. Rosemary Grant. 2002. Unpredictable Evolution in a 30-Year Study of Darwin’s Finches. Science vol 296: 707-711.
↑ McNew et al.. 2017. Epigenetic variation between urban and rural populations of Darwin’s finches. BMC Evol Biol 17, 183. DOI: 10.1186/s12862-017-1025-9
L'Escargot des haies ↑

John W. Taylor. 1894. Monograph of the land and freshwater Mollusca of the British Isles. Volumes 1 - 3. Taylor Brothers, Leeds. domaine public.
L'Escargot des haies (Cepaea nemoralis) est tout aussi célèbre que les exemples précédents; il est cité presqu'à chaque fois qu'on cherche à illustrer la variabilité naturelle (le polymorphisme). La coquille présente de grandes variations de couleur ainsi que dans la position et le nombre de bandes sombres. L'espèce se confond facilement avec sa cousine l'Escargot des jardins (Cepaea hortensis), la seule différence étant un bord (presque toujours) plus sombre. Les deux espèces s'accouplent très rarement entre elles et les descendants sont très peu fertiles.
Une sélection par la prédation
Un des prédateurs est la Grive musicienne (lorsqu'elle est présente); cet oiseau casse les coquilles sur des pierres, consomme les escargots sur place, et abandonne les coquilles vides au voisinage immédiat. Des relevés ont été réalisés pour comparer les colorations des coquilles cassées présentes au voisinage d'une "pierre à Grives" et celles des escargots vivants, dans la même zone (Cain, 1953).| escargots | sans bandes | sans bandes supérieures |
avec bandes supérieures |
|---|---|---|---|
| vivants | 153 (42%) | 143 (42%) | 264 (48%) |
| consommés | 204 (58%) | 173 (58%) | 486 (52%) |
| total | 357 | 316 | 750 |

Diagramme de corrélation entre la proportion de coquilles jaunes, la proportion de coquilles sans bandes (ou dont les bandes sont masquées) et l'habitat d'après Cain et Sheppard, 1953, fig.1
Une sélection par la température et l'humidité
Plus tard Heath et de nombreux autres auteurs (Heath, 1975) ont associé l'ensoleillement aux variations d'aspect de la coquille.Zuzanna Rosin et ses collaborateurs (Rosin, 2018) ont mené une étude très complète basée sur des élevages en laboratoire et des observations sur le terrain.
Laurence Cook (Cook, 1998) a essayé une synthèse des multiples études qui prises individuellement, n'ont pas vraiment permis de dégager une explication faisant consensus. Elle conclut que la situation actuelle résulte d'une sélection pendant des milliers d'années, le polymorphisme résultant de l'adaptation à des milieux variés sous des pressions de sélection (par la prédation, par le climat) dont les effets s'opposent. Aujourdhui l'évolution rapide des environnements conduit a une sélection et des fluctuations globalement aléatoires.
De plus, le comportement des prédateurs peut s'adapter à l'abondance des escargots, ces derniers étant délaissés pour d'autres proies lorsqu'ils sont rares. De plus le comportement des escargots dépend dans une certaine mesure de la couleur de la coquille: les escargots à coquille claire sont plus actifs en situation ensoleillée que leurs congénères à coquille sombre. Les échanges génétiques entre populations voisines mais occupant des milieux différents peuvent aussi contribuer à maintenir le polymorphisme.
Pour le profane l'exemple déconcerte puisqu'on pourrait en conclure qu'il n'y a pas d'évolution ou comme l'on fait un temps certains scientifiques, que la polymorphie de l'Escargot des haies s'explique par des variations neutres (sur lesquelles la sélection naturelle n'agit pas), ce qui est en fait inexact).
↑ A. J. Cain, P. M. Sheppard. 1953. Natural selection in Cepea. Genetics 39.
↑ D. J. Heath. 1975. Colour, sunlight and internal temperatures in the land-snail Cepaea nemoralis. Oecologia 19: 29–38. DOI: 10.1007/BF00377587
La température internes de coquilles de couleurs variées exposées au soleil est mesurée; mais c'est uniquement une expérimentation de laboratoire.
↑ Laurence Cook. 1967. The genetics of Cepaea nemoralis. Heredity 22(3). DOI: 10.1038/hdy.1967.49
↑ Laurence Cook. 1998.A two-stage model for Cepaea polymorphism. Phil. Trans. R. Soc. Lond. B 353: 1577-1593. ResearchGate.
Un bilan de toutes les hypothèses et études concernant le polymorphisme dans le genre Cepea.
↑ Laurence Cook. 2005. Disequilibrium in some Cepaea populations. Heredity 94: 497-500. DOI:10.1038/sj.hdy.6800645
↑ Laurence Cook. 2008. Variation with habitat in Cepaea nemoralis: The Cain & Sheppard diagram. Journal of Molluscan Studies 74(3). DOI:10.1093/mollus/eyn011 ResearchGate
↑ Laurence Cook. 2013. Morph frequency in British Cepaea nemoralis: What has changed in half a century ? Journal of Molluscan Studies 80(1):43-46. DOI:10.1093/mollus/eyt040. ResearchGate.
↑ . Schilthuizen. 2013. Rapid, habitat-related evolution of land snail colour morphs on reclaimed land. Heredity 110: 247–252. DOI: 10.1038/hdy.2012.74
↑ Zuzanna M. Rosin et al.. 2018. Shell colour, temperature, (micro)habitat structure and predator pressure affect the behaviour of Cepaea nemoralis. Naturwissenschaften 105(5-6):35, DOI: 10.1007/s00114-018-1560-2 Pubmed
↑ D. J. Heath. 1975. Colour, sunlight and internal temperatures in the land-snail Cepaea nemoralis. Oecologia 19: 29–38. DOI: 10.1007/BF00377587
La température internes de coquilles de couleurs variées exposées au soleil est mesurée; mais c'est uniquement une expérimentation de laboratoire.
↑ Laurence Cook. 1967. The genetics of Cepaea nemoralis. Heredity 22(3). DOI: 10.1038/hdy.1967.49
↑ Laurence Cook. 1998.A two-stage model for Cepaea polymorphism. Phil. Trans. R. Soc. Lond. B 353: 1577-1593. ResearchGate.
Un bilan de toutes les hypothèses et études concernant le polymorphisme dans le genre Cepea.
↑ Laurence Cook. 2005. Disequilibrium in some Cepaea populations. Heredity 94: 497-500. DOI:10.1038/sj.hdy.6800645
↑ Laurence Cook. 2008. Variation with habitat in Cepaea nemoralis: The Cain & Sheppard diagram. Journal of Molluscan Studies 74(3). DOI:10.1093/mollus/eyn011 ResearchGate
↑ Laurence Cook. 2013. Morph frequency in British Cepaea nemoralis: What has changed in half a century ? Journal of Molluscan Studies 80(1):43-46. DOI:10.1093/mollus/eyt040. ResearchGate.
↑ . Schilthuizen. 2013. Rapid, habitat-related evolution of land snail colour morphs on reclaimed land. Heredity 110: 247–252. DOI: 10.1038/hdy.2012.74
↑ Zuzanna M. Rosin et al.. 2018. Shell colour, temperature, (micro)habitat structure and predator pressure affect the behaviour of Cepaea nemoralis. Naturwissenschaften 105(5-6):35, DOI: 10.1007/s00114-018-1560-2 Pubmed
Avec l'œil des généticiens ↑
La génétique de Biston betularia est en fait assez mal connue. Le temps de génération calé sur les saisons est de un an ce qui nécessite d'être patient. Au début du 20e siècle, W. Bowater (Bowater, 1914), par la pratique classique (mais assez nouvelle à une époque où on redécouvrait le travail de Mendel) de croisement a étudié la transmission du mélanisme chez plusieurs espèces de papillons; malheureusement sa publication, bien qu'ancienne n'est pas en Open access; il ne semble pas avoir étudié B. betularia, mais au moins une espèce présentant une mutation du même type (Odontopera bidentata).Hors si on sélectionne correctement les formes carbonaria sur plusieurs générations pour disposer d'une variété stable dans sa couleur, un croisement carbonaria x typica donne des hybrides d'aspect carbonaria et les hybrides croisés avec typica donnent une ségrégation mendelienne (50% de chaque phénotype). On peut en déduire que carbonaria diffère de typica par une mutation sur un locus unique, et que cette mutation est dominante.
Mais cela n'explique pas les formes intermédiaires nommées (en Angleterre) insularia. Il faut donc faire intervenir l'existence de plus de deux allèles sur ce locus ou l'intervention de mutations sur d'autres gènes (ou les deux.)
Kettlewell avait montré en 1965 que certaines souches pouvaient annuler la dominance de carbonaria.
D. Lees et E. Creed (Lees et Creed, 1977) ont réalisé des croisements de souches insularia obtenues par prélèvements en divers lieux entre elles puis avec des souches carbonaria et typica. Leur conclusion est que les phénotypes les plus proches (très sombres ou très clairs) dépendent de la mutation d'un locus identique à celui expliquant le phénotype carbonaria, mais que les les phénotypes les plus intermédiaires dépendraient d'autres gènes, les résultats n'étant pas interprétables dans le cadre d'une ségrégation mendélienne, ce qui confirme les affirmations de Kettlewell (1955).
Chez la Phalène, l'insertion d'un transposon dans un gène régulateur explique l'évolution des phénotypes
On sait aujourd'hui (van't Hof, 2016) que la différence entre les deux principales variétés de Phalènes est due à l'insertion chez carbonaria d'un transposon au niveau du gène cortex (une insertion de près de 22 000 nucléotides) situé sur le chromosome 17; la suite n'est pas connue avec précision mais le gène cortex (qui code une protéine) régulerait la prolifération cellulaire en particulier dans les disques imaginaux à l'origine des ailes (il pourrait avoir d'autres rôles); reste à expliquer le changement de la couleur du corps. Sur la base de la comparaison de marqueurs du locus cortex de nombreux papillons de la variété carbonaria, van't Hof estime que l'insertion résulte d'un événement unique et daté du début de 19e siècle ( van't Hof, 2011). Les gènes souvent impliqués dans le mélanisme des insectes ne montrent eux aucune différence entre les deux variétés carbonaria et typica (Arjen van't Hof, 2010). Une hypothèse envisageable est qu'au moins certains de ces gènes fonctionneraient en réseau et que leur expression serait modulée par le gène cortex ?En 2019 Arien van't Hof et ses collaborateurs (van't Hof, 2019) on étudié des marqueurs du locus cortex chez deux espèces de Papillons autres que la Phalène présentant un mélanisme industriel. Ce même locus serait concerné mais s'il s'agissait à nouveau de l'insertion d'un transposon, elle ne serait pas au même endroit; et il pourrait tout aussi bien s'agir de la modification d'une zone régulatrice ou du gène cortex lui-même changeant la séquence de sa protéine; cette mutation serait plus ancienne que chez B. betularia (avant le début de la période industrielle).
Heliconius: ce genre de papillon montre de grandes variations reliées à plusieurs gènes dont le gène cortex, mais aussi à d'autres.
Si on découvrait d'autres exemples (de mutations liées au déplacement d'un transposon), la Phalène pourrait être une bonne illustration du rôle des transposons dans l'apparition de certaines mutations.
↑ W. Bowater. 1914. Heredity of melanism in Lepidoptera. J Genet 3: 299–315.
↑ D. R. Lees & E. R. Creed. 1977. The Genetics of the insularia form of the Peppered Moth. Heredity 39(1): 67-73.
↑ Vincent Guili. 2016. La mutation à l'origine du mélanisme industriel de la Phalène du bouleau. Planet Vie ENS.
↑ Arien E. vant'Hof et al. 2016. The industrial melanism mutation in British peppered moths is a transposable element. Nature 534: 102–105. GOI:10.1038/nature17951 core.ac.uk
↑ Arien E. van't Hof et al.. 2019. Genetic convergence of industrial melanism in three geometrid moths. Biol Lett. 15(10):20190582, DOI: 10.1098/rsbl.2019.0582 PubMed
↑ D. R. Lees & E. R. Creed. 1977. The Genetics of the insularia form of the Peppered Moth. Heredity 39(1): 67-73.
↑ Vincent Guili. 2016. La mutation à l'origine du mélanisme industriel de la Phalène du bouleau. Planet Vie ENS.
↑ Arien E. vant'Hof et al. 2016. The industrial melanism mutation in British peppered moths is a transposable element. Nature 534: 102–105. GOI:10.1038/nature17951 core.ac.uk
↑ Arien E. van't Hof et al.. 2019. Genetic convergence of industrial melanism in three geometrid moths. Biol Lett. 15(10):20190582, DOI: 10.1098/rsbl.2019.0582 PubMed
Des supergènes coderaient la forme du bec et la taille chez les Pinsons de Darwin
Erik Enbody et son équipe (Entbody, 2023) ont séquencé la totalité du génome de 4 000 pinsons présents sur Daphne et appartenant à 4 espèces (2023). Six locus (qui peuvent correspondre chacun à plusieurs gènes), tous situés sur le chromosome 1A semblent jouer un rôle majeur dans la variation; ces six locus expliquent 45% de la variation de la taille du bec de G. fortis.Le locus G03 expliquerait à lui seul 25% de la variation de la taille du bec et 13% de la variation de la taille du corps (G03 regroupe 4 gènes (dont le gène HMGA2) contrôlant ces deux "caractères" (Lamichhaney, 2016). G03 serait un supergène impliquant mutations et interactions épistatiques.
Le locus G07, proche de G03, est représentatif de 14% de la variation chez G. fortis. Un grand nombre de mutations faux-sens sont reliées au locus G07; deux concernent le gène codant le facteur de transcription ALX1 connu par ailleurs (Lamichhaney, 2015) et 4 au gène LRRIQ1 possédant un domaine de liaison avec la calmoduline; le locus G07 pourrait aussi être un supergène.
La protéine BMP4 associée à des variations dans la morphologie du bec (Abzhanov, 2004) (Abzhanov, 2006) (Florent Campo-Paysaa. 2008.) n'apparait reliée à aucun QTL (Quantitative Trait Loci). Les groupes de gènes (supergènes) apparaitraient dans l'adaptation à l'environnement et joueraient un rôle important dans la radiation adaptative. L'existence d'échanges génétiques entre espèces/variétés peut en complément de la sélection par l'environnement avoir de grandes conséquences dans une organisation du génome implicant des supergènes.
En 2018 Sangeet Lamichhaney et ses collaborateurs (Lamichhaney, 2018) ont révélé ce qui semble être l'apparition d'une nouvelle espèce de Pinson sur l'ile de Daphne. hors cette spéciation s'est réalisée en 3 générations...
↑ Arhat Abzhanov et al.. 2004. Bmp4 and morphological variation of beaks in Darwin's finches. Science 305 (5689) : 1462-5.
↑ Arhat Abzhanov et al.. 2006. The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches. Nature 442: 563-567.
F↑ Florent Campo-Paysaa. 2008. Sélection naturelle: ce que Darwin ne pouvait pas savoir. Planet-terre ENS.
↑ Ricardo Mallarino et al.. 2011. Two developmental modules establish 3D beak-shape variation in Darwin’s finches. Proceedings of the National Academy of Sciences 108(10): 4057-62. DOI:10.1073/pnas.1011480108
↑ Sangeet Lamichhaney et al.. 2015. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature volume 518: 371–375.
↑ Sangeet Lamichhaney et al.. 2016. A beak size locus in Darwin’s finches facilitated character displacement during a drought. Science vol 352 Issue 6284: 470-473. ResearchGate.
↑ Fan Han et al.. 2017. Gene flow, ancient polymorphism, and ecological adaptation shape the genomic landscape of divergence among Darwin's finches. Genome Res. 2017 27(6): 1004-1015. doi: 10.1101/gr.212522.116
↑ Sangeet Lamichhaney et al.. 2018. Rapid hybrid speciation in Darwin’s finches. Science 359 ( 6372): 224-228. DOI:10.1126/science.aao4593
↑ Erik D. Enbody et al.. 2023. Community-wide genome sequencing reveals 30 years of Darwin’s finch evolution. Science 381 eadf6218. DOI:10.1126/science.adf6218
↑ Arhat Abzhanov et al.. 2006. The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches. Nature 442: 563-567.
F↑ Florent Campo-Paysaa. 2008. Sélection naturelle: ce que Darwin ne pouvait pas savoir. Planet-terre ENS.
↑ Ricardo Mallarino et al.. 2011. Two developmental modules establish 3D beak-shape variation in Darwin’s finches. Proceedings of the National Academy of Sciences 108(10): 4057-62. DOI:10.1073/pnas.1011480108
↑ Sangeet Lamichhaney et al.. 2015. Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature volume 518: 371–375.
↑ Sangeet Lamichhaney et al.. 2016. A beak size locus in Darwin’s finches facilitated character displacement during a drought. Science vol 352 Issue 6284: 470-473. ResearchGate.
↑ Fan Han et al.. 2017. Gene flow, ancient polymorphism, and ecological adaptation shape the genomic landscape of divergence among Darwin's finches. Genome Res. 2017 27(6): 1004-1015. doi: 10.1101/gr.212522.116
↑ Sangeet Lamichhaney et al.. 2018. Rapid hybrid speciation in Darwin’s finches. Science 359 ( 6372): 224-228. DOI:10.1126/science.aao4593
↑ Erik D. Enbody et al.. 2023. Community-wide genome sequencing reveals 30 years of Darwin’s finch evolution. Science 381 eadf6218. DOI:10.1126/science.adf6218
L'Escargot des haies
Des études par croisement ont été menées dès le début du 20e siècle.
Les gènes déterminant la couleur de la coquille et la formation des bandes sont fortement liés (le taux de recombinaison avoisine 1%) (Cook et King, 1966). Un groupe de gènes indépendants du précédent modifie le nombre de bandes (ils ne s'expliment que chez un homozygote pour la formation des bandes) et quelques autres locus sont également impliqués.Sur court terme un déséquilibre entre des pressions de sélection différentes (visuelles et non visuelles) explique, avec la migration d'individus d'un habitat à un autre, le maintien du polymorphisme. Sur le long terme il semble que la sélection visuelle ait conduit à rapprocher des gènes pour verrouiller les recombinaisons alléliques. On n'explique cependant pas pourquoi le contrôle des motifs formés par les bandes et la couleur dépendent de groupes différents.
↑
D. R. Gonzalez et al.. 2019. Recombination within the Cepaea nemoralis supergene is confounded by incomplete penetrance and epistasis. Heredity 123: 153–161. DOI: 10.1038/s41437-019-0190-6
La recombinaison au sein du supergène de Cepaea nemoralis est masquée par une pénétrance (expression) incomplète et une épistasie (interaction entre gènes).
↑ Margrethe Johansen. 2023. Fine mapping of the Cepaea nemoralis shell colour and mid-banded loci using a high-density linkage map. Heredity 131(5-6): 327–337. DOI: 10.1038/s41437-023-00648-z
La recombinaison au sein du supergène de Cepaea nemoralis est masquée par une pénétrance (expression) incomplète et une épistasie (interaction entre gènes).
↑ Margrethe Johansen. 2023. Fine mapping of the Cepaea nemoralis shell colour and mid-banded loci using a high-density linkage map. Heredity 131(5-6): 327–337. DOI: 10.1038/s41437-023-00648-z
Perspectives pour l'enseignement (détails) ↑
Trop beau pour être vrai
On pourrait penser que l'exemple de la Phalène est d'une simplicité caricaturale et qu'il est, pour cette raison, un piètre choix pour mettre en évidence le polymorphisme, en contradiction avec l'idée que la variabilité naturelle des populations est le fondement du modèle darwinien de l'évolution. L'objectivité doit (malheureusement ?) nous conduire à reconnaitre qu'on l'a rendu caricatural. 
Fréquence de la variété carbonaria autour de Manchester. Les relevés (points noirs -les barres verticales représentent la marge d'erreur-) sont comparés à une courbe calculée à l'aide du modèle de Cook et Turner (2008):
q' = q[ 1-s(1-q)]/[1-s(1-q2)]
avec q fréquence de typica, q' sa fréquence à la génération suivante (un an plus tard), s le coefficient de sélection de carbonaria en lui donnant une valeur de 25%; d'après Cook, 2011, fig.1Vue de cette manière la variabilité n'existe que dans le temps.
Le "roman" de l'évolution
Et pourtant alors les Phalènes entièrement noires disparaissent, il existe encore (et même de plus en plus) des Phalènes partiellement noires, celles dénommées insularia en Angleterre. Leur existence est totalement escamotée par le récit (en fait le roman) que font les manuels de l'histoire de la Phalène. Cette transposition didactique est difficilement acceptable (heureusement les créationistes qui ont critiqué l'expérience se sont focalisés sur des détails ridicules comme la relation entre les troncs et le comportement des Phalènes et ne sont pas allés lire l'article original de Kettlewell).Cette analyse ne retire absolument rien à la valeur scientifique des expérimentations de Kettlewell, d'autant Kettlewell prend en compte ces formes intermédiaires dans son protocole et a suggéré par ailleurs que plusieurs mutations et plusieurs gènes étaient en jeu dans la coloration des ailes ou du corps.
L'étude des manuels scolaires (présente sur la page détaillée) montre que nombre de points essentiels sont maltraités ou non traités:
- la non induction du mélanisme (au niveau génétique) par le changement de l'environnement (en fait difficile à démontrer dans la mesure ou la mutation semble résulter d'un évenement fortuit unique postérieur au changement d'environnement);
- l'héritabilité du caractère (un seul manuel évoque la réalisation de croisements sans fournir de détails, les autres utilisent l'argument d'autorité: l'égalité un caractère = un gène est affirmée sans justification (alors qu'on sait qu'elles est en grande partie fausse).
- l'existence de formes intermédiaires certes peu fréquentes est escamotée. Pourtant, cela retirerait ne rien à la pertinence de l'expérimentation de Kettlewell. On joue sur de petits effectifs mais les résultats sont intermédiaires, ce qui confirme plutôt les résultats obtenus pour les phénotypes les plus fréquents. Les mentionner montre que la coloration des ailes est moins simple qu'il n'y parait et que l'idée un caractère = un gène induite par la simplification est totalement fausse, une situation qui gène beaucoup les réductionnistes.
Cet exemple illustre au moins la rapidité avec laquelle des espèces peuvent s'adapter sans que cela repose forcément sur une variabilité initiale; il faut surtout signaler qu'il est regrettable de généraliser à partir d'un exemple unique (et tronqué) sous prétexte qu'il est "plus simple".
L'expérience de Kettlewell. Wikiland.
Une description assez précise de l'expérience suivie d'un imposant développement sur les critiques des créationnistes américains.
↑ Laurence Cook & J. Saccheri. 2013. The peppered moth and industrial melanism: evolution of a natural selection case study. Heredity 110: 207–212. DOI: hdy.2012.9243-46
↑ Laurence Cook. 2011. The melanic form of the peppered moth, Biston betularia, in Manchester: the end of an era. Entomologist s Gazette 62(2):91-99. ReseachGate.
↑ Laurence M. Cook and JRG Turner. 2008. Decline of melanism in two British moths: spatial, temporal and inter-specific variation. Heredity 101: 483–489.
Une description assez précise de l'expérience suivie d'un imposant développement sur les critiques des créationnistes américains.
↑ Laurence Cook & J. Saccheri. 2013. The peppered moth and industrial melanism: evolution of a natural selection case study. Heredity 110: 207–212. DOI: hdy.2012.9243-46
↑ Laurence Cook. 2011. The melanic form of the peppered moth, Biston betularia, in Manchester: the end of an era. Entomologist s Gazette 62(2):91-99. ReseachGate.
↑ Laurence M. Cook and JRG Turner. 2008. Decline of melanism in two British moths: spatial, temporal and inter-specific variation. Heredity 101: 483–489.
Les supergènes jouent-ils un rôle crucial ?
Les deux autres exemples ont étés choisis en raison de leur présence dans les manuels ou dans des suggestions d'activités didactiques. Dans les deux cas, les caractères étudiés montrent un polymorphisme important et ces caractères ont un rôle souvent crucial dans la survie. Hors on constate à chaque fois l'implication de supergènes. Ce n'est probablement pas le fait du hasard. Les supergènes sont connus depuis longtemps (chez les insectes sociaux, dans le mimétisme d'autres insectes) (Nabours, 1929), mais leur importance avait été sous-estimée, sauf par quelques-uns:"the evolution of super-genes, whether consisting of a few closely-linked loci or of an inversion, should always be (but never, I think, is) treated as one of the fundamental properties of genetics."
Edmund B. Ford. 1965. Genetic Polymorphism. Faber & Faber: London.
Ford était un spécialiste de la génétique environnementale (ecological genetics). Il a supervisé le jeune Kettlewell dans ses travaux sur la Phalène, ce qui lui a valu de nombreuses critiques et même d'être accusé de fraude (par des créationnistes), des accusations totalement injustifiées (Majerus, 2000).
La page Wikipedia conscrée aux supergènes n'existe qu'en 3 langues !
La facilité (relative) offerte par les techniques de séquençage généralisé semble aujourd'hui monter une implication des supergènes dans les mécanismes évolutifs bien plus fréquente que soupçonnée.
Enfin les supergènes n'expliquent pas tout: Chez Geospiza fortis, 6 locus dont deux représentent des supergènes expliqueraient 45% de la variation de la taille et de la forme du bec (Entbody, 2023); restent quand même 55% a expliquer par ailleurs... Il se faut se garder de toute attitude réductionniste.
Par rapport aux exemples qui précèdent l'escargot a un avantage, c'est que le polymorphisme, sauf en milieu très urbanisé est facilement observable, et un inconvénient: peu de données existent montrant une évolution en un lieu précis.
↑ Supergene. Wikipedia (en).
↑ M. N. Majerus et al.. 2000. A bird's eye view of the peppered moth. Journal of Evolutionary Biology Volume 13, Issue 2. 155-159.
↑ Jim Mallet. 2003. The peppered moth: a black and white story after all. Genetics Society News, Issue 50: 34-38.
↑ M. N. Majerus et al.. 2000. A bird's eye view of the peppered moth. Journal of Evolutionary Biology Volume 13, Issue 2. 155-159.
↑ Jim Mallet. 2003. The peppered moth: a black and white story after all. Genetics Society News, Issue 50: 34-38.
Résistances ↑
Deux exemples majeurs se partagent ce chapitre: la résistance des insectes et autres arthropodes aux insecticides et la résistance des bactéries aux antibiotiques. Loin d'être anodins, ils ont une importance considérable dans le domaine économique et dans le domaine de la santé.Pour la résistance aux insecticides, l'expérience du DDT sera mise en avant: nous disposons d'un recul de temps conséquent: la première utilisation date de 1943 et l'usage en a été progressivement abandonné dans la plupart des pays (sauf l'Inde). L'utilisation du DDT répondait à plusieurs objectifs, agricoles mais surtout de santé humaine avec le contrôle de la malaria, une parasitose représentant une préoccupation majeure. Pendant plus d'une décenie l'OMS consacrera le tiers de son budget pour organiser des pulvérisations de DDT dans les foyers. Enfin, même s'il n'est pas toujours facile d'isoler la dangerosité du DDT de celle d'autres polluants organiques persistants (POP), et si on peut penser que davantage d'études auraient pu être menées sur le sujet, la littérature scientifique sur le DDT est loin d'être absente. Par ailleurs l'usage ou le non usage du DDT a suscité des débats considérables.
La Saga du DDT ↑
«Si Darwin était encore de ce monde, il serait enchanté, voire étonné, de constater à quel point le monde des insectes justifie sa théorie de la sélection naturelle. Sous les pluies d'insecticides, les membres les moins robustes de ces familles sont en train de disparaitre, et seuls demeurent pour défier nos efforts les plus puissants et les mieux adaptés.»Rachel Carlson, 1963: p.283 (chapitre 16)
Lorsqu'il est synthétisé en 1873 à Strasbourg (c'est une molécule artificielle), le DDT n'intéresse personne; Il faut attendre 1939 à Bâle, pour que la firme Geigy, en recherchant des alternatives à l'emploi de l'arsenic, le redécouvre et découvre surtout son pouvoir de destruction des Arthropodes.

Pulvérisation de poudre de DDT lors d'une épidémie de typhus, Naples, 1945; d'après une vidéo Internet Archive.

Pulvérisation d'un mélange de DDT et de kérosène pour l'élimination de la malaria, Italie, 1945; crédit: National Museum of Health and Medicine / Wikimedia, domaine public.
Le DDT devient secret défense
En 1943 l'armée américaine déclare le produit inoffensif pour les humains avant d'engager une campagne d'aspersion massive des troupes engagées en Asie (plus de la moitié des soldats étaient atteints du paludisme). Le DDT est alors considéré comme un avantage décisif sur l'armée japonaise et tout ce qui l'entoure érigée au rang de secret défense. Le DDT utilisé par les armées américaines contribue aussi à l'éradication du paludisme en Europe, même si les mesures d'hygiène et de contrôle des zones humides, le traitement des malades par la quinine en avait déjà considérablement réduit la fréquence (en Corse le paludisme disparait après 1948). On savait cependant, après des tests menés au National Institute of Health (NIH) et à la Food and Drug Administration (FDA) que le DDT à forte dose provoquait des convulsions et des troubles hépatiques pouvant conduire à la mort chez les mammifères de laboratoire. Et dès 1944, Herbert Calvery, de la FDA constate que de faibles doses accumulées au fil du temps provoquent les mêmes troubles que l’exposition à une forte dose. (Elena Conis, 2017 Science History Museum.)

Pictogramme "toxique" du système généralisé d'étiquetage des produits chimiques.
Un produit miracle ?

DDT for control of household pests. Brochure du Public Health Service, 1947.

Tirez pour tuer, protégez votre jardin de la victoire, 1943. crédit: National Archives at College Park
Les publicités de l'époque présentent le DDT comme un produit miracle et Paul Müller, l'employé de Ceigy qui l'avait redécouvert reçoit même le prix Nobel de médecine en 1948.
Une inefficacité que la lecture (et surtout la compréhension) de Darwin (L'Origine) aurait permis d'anticiper
Mais c'est l'Organisation mondiale de la santé (OMS, WHO) qui va assurer l'essentiel de la promotion. A partir de 1955 et dans la décennie qui suit, l'OMS va consacrer jusqu'à un tiers de son budget à une campagne "d'éradication du paludisme" basée sur la pulvérisation du DDT à l'intérieur des habitations, dans une "guerre contre la maladie" assimilée à la guerre tout court. L'objectif est d'une prétention incroyable; pourtant des exemples de résistance des moustiques étaient connus dès 1953 et, évidemment, le phénomène de résistance aurait été anticipé si on avait compris Darwin. Et comment ne pas considérer l'OMS d'une extraordinaire naïveté quand on pense à la complexité du cycle de développement du parasite (le Plasmodium) ? Après une baisse initiale, le paludisme augmente de nouveau en Inde et au Sri Lanka, avec l'apparition de vecteurs (moustiques Anophèles) résistants au DDT et d'une résistance du parasite lui même aux médicaments. L'échec conduit l'OMS à revoir sa stratégie en 1969.Un polluant organique persistant
Le DDT est une molécule organo-chlorée très stable dont la demi-vie est évaluée entre 2 et 15 ans. Comme il n'est pas ou très peu métabolisé par les organismes il se concentre au long des chaines alimentaires, ce qui augmente sa dangerosité pour les prédateurs. 
© Concentration en DDT le long des chaines alimentaires, Long Island, New York, 1968. crédit: National Museum of Health and Medicine / Wikipedia
Le DDT est considéré comme très dangereux pour les arthropodes aquatiques. Il provoque un amincissement des la coquille des oiseaux. Son action de perturbateur endocrinien est bien établie, y compris chez les humains. Par ailleurs les produits de dégradation (DDE) sont eux même très stables et toxiques. La dangerosité des deux molécules pour les humains a longtemps été peu étudiée (Eskenazi, 2009)..

Dans de nombreux pays, une baisse initiale de l'incidence du paludisme est suivie de remontées exponentielles. Le lobby pro DDT a accusé les détracteurs d'être à l'origine des échecs, ce qui évitait de s'interroger sur les défauts des procédures. Hors, l'incidence de la maladie a été multipliée par trois entre 1961 et 1966 en Inde (et dans de nombreux autres pays), 6 ans avant l'interdiction du DDT aux USA. Le phénomène d'apparition de résistance a été systématiquement occulté. Même si des études ultérieures ont estimé que des défauts d'organisation étaient responsables dans 2/3 des cas d'échecs, et la sélection de moustiques résistants dans seulement 1/3, ce constat aurait mérité des analyses plus approfondies, en particulier pour suivre l'apparition de résistances.
L'OMS va persister à défendre les pulvérisations de DDT dans les locaux d'habitation jusqu'en 2006. En 2008, de nombreux chercheurs réunis lors d'une conférence compilent les études disponibles et signent "The Pine River Statement"; ils soulignent les risques connus, recommandent de n'utiliser le DDT qu'en dernier recours et réclament des études complémentaires concernant les effets sur la santé humaine (Eskenazi, 2009). D'autres approches délaissant le DDT ont été menées ensuite, en mettant en jeu simultanément plusieurs barrières (moustiquaires imprégnées, traitement des malades). Inspirés par l'élimination du paludisme au Sri Lanka, l'Inde vise actuellement le même but en se focalisant sur le traitement des malades. On privilégie les traitements mixtes mais surtout.
↑ et al.. 2009. doi: 10.1289/ehp.11748. NIH.
↑ B. Eskenazi B et al.. 2009. The Pine River statement: human health consequences of DDT use. Environ Health Perspect 117(9):1359-67. doi: 10.1289/ehp.11748. NIH.
We reviewed epidemiologic studies published from 2003 to 2008 that investigated the human health consequences of DDT and/or DDE exposure.
Directives pour la prévention et la gestion de la résistance aux pesticides. FAO.
↑ B. Eskenazi B et al.. 2009. The Pine River statement: human health consequences of DDT use. Environ Health Perspect 117(9):1359-67. doi: 10.1289/ehp.11748. NIH.
We reviewed epidemiologic studies published from 2003 to 2008 that investigated the human health consequences of DDT and/or DDE exposure.
Directives pour la prévention et la gestion de la résistance aux pesticides. FAO.
Insecticides, herbicides, pesticides ↑
Un projet pharaonique
La Mission Interministérielle d’Aménagement Touristique du Littoral du Languedoc-Roussillon (plus connue comme mission Racine, du nom de son coordonnateur) est crée en 1963. Il s'agit de concurrencer l'Espagne qui développe un tourisme de masse sur les plages de la Costa Brava. L'économie locale ne repose alors que sur la culture de la vigne qui décline et les côtes qui présentent 66.000 ha de zones humides rendues inhospitalières par les moustiques.Vivre avec les moustiques
L'objectif de la mission se veut réaliste dès le départ: il s'agit de réduire la nuisance des moustiques et non de viser l'éradication, d'autant qu'aucune maladie grave n'est à l'époque potentiellement transmise par ces moustiques. Tout va reposer sur l'Entente interdépartementale de démoustication (EID). Cet organisme est peu disert sur les méthodes utilisées à l'origine qui consistaient à des épandages massifs de DDT, souvent par avion, puis d'organophosphorés comme le fénitrothion. Le fénitrothion, bien moins persistant que le DDT sera utilisé ultérieurement dans l'ile de la Réunion. L'EID est très disert sur le Bti (Bacillus thuringiensis ser. israelensis), une bactérie agissant comme biocide ciblé sur les larves en milieu aqueux, complété par la deltaméthrine, issue de pyrèthres naturels. Les propriétés du Bti ont été découvertes en 1976, sa fabrication est un monopole de Valent Biosciences, une société américaine et les adjuvents constituent un secret commercial, mais l'EID le présente comme un produit miracle.
Lorsque les spores de Bti sont ingérées par les larves de diptères, en particulier par les larves de moustiques, la digestion produit 4 toxines qui perforent la paroi intestinale des larves. Selon l'EID aucune résistance n'a été observée après plusieurs décenies d'utilisation. Cependant le Bti n'est actif que sur les 3 premiers stades larvaires des moustiques (qui en comptent 4) et il doit être avalé par la larve dans les 24 heures après avoir été répandu dans le milieu. De plus le Bti détruit aussi les larves de Chironomes et on l'accuse ainsi de perturber l'écosystème lagunaire (V, 2020). On considère que 80% des larves sont détruites lors d'un traitement, ce qui n'élimine pas totalement les moustiques (au grand regret de l'EID), les opérations doivent être renouvellées chaque année et nécessitent un suivi écologique précis des gites larvaires des lagunes pour bien choisir le lieu et le moment de l’épandage.
L'usage de la deltaméthrine (disponible aussi en grandes surfaces) concerne plutôt les milieux urbains. Une seule espèce de moustique a longtemps été présente dans ces milieux: Culex pipiens. Malheureusement Aedes albopictus ou Moustique tigre s'y est ajoutée depuis peu. Les oeufs du Moustique tigre sont réputés avoir une longue durée de survie, y compris pendant l'hiver. Ce moustique pique plutôt à l'extérieur en début et en fin de journée et les larves affectionnent les petits récipients d'eau claire (soucoupes de pots de fleurs, eau retenue dans les grandes feuilles, etc) L'usage de la deltaméthrine devrait être plus contrôlé pour éviter l'apparition de résistances. Le Moustique tigre peut être vecteur de nombreux virus, les plus préoccupants étant ceux de la dengue et du chikungunya. La participation de la population locale semble indispensable dans l'élimination des gites permettant l'éclosion des œufs, hors certaines enquêtes montrent que les représentations sont inadaptées (Mieulet, 2016). Malgré cette inadaptation, certaines préparations de Bti sont disponibles pour l'usage domestique et agréées pour l'agriculture biologique (Terra nostra).

Les baraques à Gruissan, début 20e siècle.

Gruissan: la plage, début 20e siècle.
Pour être complet, si les villages languedociens étaient tous éloignés de la mer de 1 à 3 km, les moustiques n'étaient pas seuls responsables et n'étaient même pas aussi dissuasifs qu'il a été dit pour les amateurs de plages:
- par l'éloignement, on se protégeait des coups de mer sur un littoral très bas;
- il existait des habitats précaires construits en bois ou en roseau et plus sagement sur pilotis comme les cabanes de pêcheurs de Gruissan. Et leurs occupants temporaires s’accommodaient des moustiques dès la fin du 19e siècle...

Gruissan le 25 novembre 2021; © ville de Gruissan.
L'aménagement du littoral languedocien survivra-t-il au réchauffement climatique ?
On peut estimer que la mission Racine est un succès en ce qui concerne le contrôle des moustiques et même en ce qui concerne l'aménagement du littoral, mais seulement à court terme. Sur le long terme, se prémunir de l'apparition de souches résistantes de moustiques reste une grande préoccupation et plus grave, même si on sort un peu du sujet de cette page, le recul du trait de côte du à la modification des courants marins et à la baisse des alluvions apportées par les rivières est dramatique, sans compter l'élévation prévue du niveau de la mer. A Gruissan la mission Racine s'est sagement inspirée de la tradition en conservant les constructions sur pilotis, mais on envisage... d'expérimenter des habitats flottants.
↑ Tout savoir sur le Bti. EID-méditerranée.
↑ Brühl C.A. et al.. 2020. Environmental and socioeconomic effects of mosquito control in Europe using the biocide Bacillus thuringiensis subsp. israelensis (Bti). Science of the Total Environment 724: 137800. DOI: 10.1016/j.scitotenv.2020.137800
↑ Elise Mieulet et Cécilia Claeys. 2016. (In)acceptabilités environnementales et/ou sanitaires : dilemmes autour de la démoustication du littoral méditerranéen français. VertigO Volume 16 Numéro 1. DOI : https://doi.org/10.4000/vertigo.16940
↑ Anti Moustiques Larvicides.
En 1965, après le développement de la résistance au DDT chez le Pou, l'armée américaine le remplace par le lindane HCH) dans la lutte contre le typhus (il est toujours autorisé aujourd'hui aux Etats-Unis dans le domaine médical, mais en seconde intention, quand d'autres traitements ont échoué).↑ Brühl C.A. et al.. 2020. Environmental and socioeconomic effects of mosquito control in Europe using the biocide Bacillus thuringiensis subsp. israelensis (Bti). Science of the Total Environment 724: 137800. DOI: 10.1016/j.scitotenv.2020.137800
↑ Elise Mieulet et Cécilia Claeys. 2016. (In)acceptabilités environnementales et/ou sanitaires : dilemmes autour de la démoustication du littoral méditerranéen français. VertigO Volume 16 Numéro 1. DOI : https://doi.org/10.4000/vertigo.16940
↑ Anti Moustiques Larvicides.
Une fuite en avant qui revient à nier la sélection naturelle tant qu'elle ne diminue pas les profits
De nombreux insecticides figurent dans la liste des POP (Polluant Organique Persistant) établie en 2004 par la Convention de Stockholm comme le DDT, la dieldrine (interdite en France en 1972), le lindane, le chlordécone (interdit aux Etats-Unis en 1978 et aux Antilles françaises seulement en 1993)... Le chlordécone considéré comme non dégradable a une demi-vie évaluée à 650 ans dans le sol et est à l'origine d'un scandale sanitaire en France en raison des délais accordés avant l'interdiction de son usage massif dans les cultures de bananes; il est fortement cancérigène.L'effet insecticide des pyréthrines présentes dans les fleurs de Pyrèthre est connu depuis 2500 ans en Perse. Des pyréthrinoïdes synthétiques ont été créés dans les années 1960 pour augmenter leur stabilité; après avoir été abandonnées pour les insecticides persistants (POP), ces molécules connaissent un regain d'intérêt en raison de leur moindre toxicité. Comme le DDT elles interagissent avec le fonctionnement des canaux sodium des neurones. La grande différence avec le DDT est une biodégrabilité très forte; elles sont aussi beaucoup mieux supportées par les Oiseaux et les Mammifères. Le phénomène de résistance se manifeste sans surprise mais peut être contourné ou différé par l'usage de Piperonyl butoxide (PBO), un composé organique dérivé de substances présentes dans le Safran. Les pyréthrines, de plus mortelles pour les abeilles, ne sont donc pas la panacée, seulement un moindre mal.

fig2 p.82 Rapport de l'IGF, 2021.
En 2015, devant la faiblesse des résultats, un plan Écophyto 2 a été défini. L’objectif de -50% est repoussé en 2025 avec un objectif intermédiaire de -25% à horizon 2020.
En 2018, le Gouvernement décide un "plan d’action" sur les produits phytopharmaceutiques (PPP) et une agriculture "moins dépendante aux pesticides" complété de l’objectif de "sortir du glyphosate", et amende le plan qui prend le nom d'Écophyto 2+.
Repoussant les échéances et de diminuant les ambitions, on ne peut que constater l'échec dans la réduction de l'usage des pesticides, dénoncé dans un rapport de l'Inspection Générale des Finances de 2021 (IGF, 2021). Les plans se sont limités à mettre en valeur des bonnes pratiques, restées marginales par rapport aux pratiques majoritaires restées inchangées. Le rapport suggère d'établir des objectifs contraignants de baisse par exploitation, avec, par exemple, un système de bonus-malus.
Le comble du cynisme ou de l'absurde est atteint quand l'Europe (France comprise) exporte des pesticides dont l'usage est interdit en Europe (en particulier vers les Etats-Unis et le Brésil, vers l'Ukraine et vers le Japon pour la France); ces produits sont utilisés pour des cultures qui reviennent en Europe et en France sous forme de produits importés, toxiques pour les consommateurs et constituant une concurrence inéquitable pour nos agriculteurs.
On constate que l'usage agricole des pesticides n'a pas diminué; que la proportion des récoltes perdue à cause des insectes augmente. Souvent on attend pour abandonner un insecticide qu'il se révèle moins efficace (à cause de l'apparition de résistances et parce qu'on peut passer à un autre, qui n'est pas forcément moins nocif). Le lobby des pesticides (Reporterre, 2023) répète une histoire déjà connue, celle du tabac, celle de l'amiante...
↑ 2021. Évaluation des actions financières du programme Écophyto. Rapport de l'Inspection générale des Finances.
↑ . La carte de l'intensité d'usage des pesticides dans votre intercommunalité (France).
L'ensemble de ce site Résilience alimentaire (soutenu par l'ADEME et le Ministère de l'agriculture (!) est très intéressant par ailleurs.
↑ . 2021. Pesticides et santé – Nouvelles données. INSERM.
↑ . 2021. Le lobby des pesticides accusé de chantage à l’emploi mensonger. Reporterre.
↑ . La carte de l'intensité d'usage des pesticides dans votre intercommunalité (France).
L'ensemble de ce site Résilience alimentaire (soutenu par l'ADEME et le Ministère de l'agriculture (!) est très intéressant par ailleurs.
↑ . 2021. Pesticides et santé – Nouvelles données. INSERM.
↑ . 2021. Le lobby des pesticides accusé de chantage à l’emploi mensonger. Reporterre.
Les antibiotiques, c'est pas automatique ↑
Expérimenter avec élégance
A la suite de Salvador Luria et Max Delbrück qui en 1943 mettent en culture des bactéries et des phages (virus), puis de Newcombe (Newvcombe, 1949), Esther & Joshua Lederberg (Lederberg, 1952) expérimentent sur la résistance de bactéries aux phages et aux antibiotiques; une technique, celle de la copie d'une culture en boite de Pétri grâce à un tampon de velours mise au point par Esther révolutionne les tests de phénotypes bactériens. Grâce à cette technique l'une de leurs expériences est particulièrement élégante et son résultat facile à comprendre. 
Le tampon de replication permet de copier le contenu de la boite maitre et de distinguer, après incubation, les colonies résistantes (à l'antibiotique) présentes dans toutes les boites et les colonies sensibles, absentes des boites avec antibiotique. Cliquez sur l'image pour la version
.svg (fichier présent sur Wikimedia, CC by SA 4.0 attribution: Michel Racine).Si la résistance était induite par l'antibiotique, les colonies résistantes apparaitraient au hasard dans les boites dont le milieu de culture est additionné d'antibiotique. Comme ce sont toujours les mêmes colonies (localisées au même emplacement sur la boite) qui sont résistantes, c'est que la résistance est apparue avant la replication. Une confirmation peut-être obtenue en repiquant sur des boites avec antibiotique des colonies sensibles (aucune bactérie ne se multiplie) et des colonies résistantes (toutes les bactéries se multiplient).
Esther & Joshua Lederberg n'ont, pas plus que Luria et Delbrück, modélisé l'évolution; Darwin en avait élaboré le modèle un siècle plus tôt; mais ils ont confimé que l'innovation précède la sélection. L'évolution n'est pas un apprentissage direct. Il est tout à fait étonnant que cette expérience, qui concerne un sujet fondamental, ne soit pas plus mise en avant par l'histoire. De plus des variantes permettent de calculer le taux d'apparition des mutations liées à un phénotype donné. Les auteurs ont montré que pour la résistance aux antibiotiques ce taux était faible, mais non nul.
Dans son manuel Génétique et évolution, Claudine Petit nous donnait un conseil qui était aussi un avertissement: «A l'intérêt théorique considérable de ces résultats s'ajoute un intérêt pratique non moins considérable (...) la mutation de plusieurs gènes assurant la résistance à des antibiotiques différents a peu de chances de se produire simultanément. De sorte que l'administration conjointe de plusieurs antibiotiques permet de neutraliser l'apparition des clones résistants.» (Petit, 1967, p. 324-325). Un conseil que nous n'avons pas suivi.
↑ Esther Lederberg & Joshua Lederberg. 1952. Replica plating and indirect selection of bacterial mutants. Journal of Bacteriology, Vol. 63, No. 3. DOI: 10.1128/jb.63.3.399-406.1952 (pdf, Pubmed)
Joshua Lederberg recevra le prix Nobel de médecine en 1958, mais pas Esther (sa découverte du phage lambda qui insère son ADN dans celui de la bactérie hôte et se multiple avec elle a pourtant fait de ce phage un organisme modèle pour une mutitude de recherches en génétique); Esther fait partie comme Rosalin Franklin des scientifiques victimes du sexisme omniprésent en recherche scientifique à l'époque. Le couple Lederberg se marie en 1946 et divorce 20 ans plus tard entrainant une ignorance quasi totale d'Esther par son ex-époux dans toutes les contacts scientifiques qui suivent leur séparation.
↑ Claudine Petit, Georges Prévost. 1967. Génétique et évolution. Hermann, Paris.
Joshua Lederberg recevra le prix Nobel de médecine en 1958, mais pas Esther (sa découverte du phage lambda qui insère son ADN dans celui de la bactérie hôte et se multiple avec elle a pourtant fait de ce phage un organisme modèle pour une mutitude de recherches en génétique); Esther fait partie comme Rosalin Franklin des scientifiques victimes du sexisme omniprésent en recherche scientifique à l'époque. Le couple Lederberg se marie en 1946 et divorce 20 ans plus tard entrainant une ignorance quasi totale d'Esther par son ex-époux dans toutes les contacts scientifiques qui suivent leur séparation.
↑ Claudine Petit, Georges Prévost. 1967. Génétique et évolution. Hermann, Paris.
Un problème de santé majeur
Une enquête réalisée par l'OMS dans de nombreux pays fait apparaitre une incompréhension totale du phénomène de résistance aux antibiotiques. Si l'idée de résistance est connue, parmi les personnes qui la connaissent, les 3/4 pensent que c'est le patient qui devient résistant et non les microbes ! Ceci empêche évidemment de comprendre qu'une personne en bonne santé pourra être victime d'une infection provoquée par une bactérie multirésistante. Presque la même proportion pense que les antibiotiques peuvent être bénéfiques pour toutes les infections (donc pour les infections virales comme le rhume et la grippe). Le rejet des élevages utilisant des antibiotiques (l'utilisation d'antibiotiques à titre préventif pour des animaux en bonne santé est interdite en Europe, mais pas ailleurs) est largement majoritaire, mais il est souvent basé sur des idées fausses. Le problème est qu'une bonne compréhension nécessite d'expliquer la résistance aux antiotiques par l'application du modèle de l'évolution darvinienne, donc la connaissance de ce modèle et son acceptation (au moins à "petite échelle").
↑ 2015. Une enquête multipays de l’OMS révèle une large incompréhension de l’opinion publique à l’égard de la résistance aux antibiotiques. OMS (WHO).
↑ Emilie Cardot Martin et al.. 2019. La résistance aux antibiotiques. Planet-vie ENS.
↑ Emilie Cardot Martin et al.. 2019. La résistance aux antibiotiques. Planet-vie ENS.
La domestication des plantes ↑
En fait les domestications, bien que les exemples servent d'arguments à Darwin dans L'Origine, restent le plus souvent des transformations limitées (du point de vue génétique). Le Maïs est presque une exception, avec des transformations morphologiques plutôt spectaculaires de la téosinte au maïs.Une race domestiquée à finalement peu de chance de survivre dans la nature (pour les plantes dont les graines sont consommées, cela peut s'expliquer en raison de la contradiction entre récolte par l'Homme et dissémination de la plante, mais cela ne vaut pas pour les autres) ce qui montre toute la limite de notre prétention. Les "améliorations" (d'un point de vue de l'utilité pour l'humanité) introduites par les sélections successives lors de la domestication le sont au détriment d'autres caractéristiques qui permettent à l'espèce de perdurer par rapport à ses concurrentes.
Une autre approche de la domestication, développée sur cette page, est d'en examiner les effets sur les écosystèmes et les sociétés humaines et cette fois la transformation touchant nos sociétés est fondamentale.
La domestication du Maïs (détails) ↑
Une seule espèce et de très nombreuses variétés
L'origine du maïs bénéficie d'une documentation considérable et d'une recherche active; l'histoire de la plante est particulièrement riche. Le Maïs cultivé Zea mays ssp mays est une graminée d’origine tropicale, de la tribu des Maydées. Différentes formes sauvages de l’espèce Zea mays existent au Mexique; ce sont les téosintes, avec des formes annuelles, Zea mays ssp mexicana et Zea mays ssp parviglumis et une forme pérenne, Zea mays ssp perennis. Si les différences morphologiques sont importantes entre les différentes formes, elles restent des variétés d'une espèce unique Zea mays. La variété perennis a subi au cours de l'histoire un doublement du nombre de ses chromosomes. Les variétés mexicana et parviglumis ont le même nombre de chromosomes que la variété cultivée (2n=10), s'hybrident facilement avec elle et la F1 issue de ces croisements est fertile.John Harshberger est le premier à avoir identifié le centre du Mexique comme origine de la plante (Harshberger, 1896: 148); Darwin l'avait déjà situé en Amérique en 1868, mais était moins précis (il insiste sur l'extraordinaire variation présentée par cette plante cultivée).

téosinte (d'après Stitzer, 2018)).

maïs cultivé (d'après Stitzer, 2018).
La comparaison maïs (cultivé) - téosinte (1) montre que:
- le port du maïs est très peu ramifié et les branches latérales se transforment en épis.
- les épis sont moins nombreux et plus gros chez le mais comparativement à la téosine.
- Le maïs possède des épillets appariés alors que le téosinte a des épillets simples (l'épillet est la petite tige portant une fleur; la plupart des herbes montrent des épillets simples, mais dans la famille du mais les épillets sont par paires dans les fleurs mâles et femelles). Dans le téosinte, bien qu'il y ait des épillets appariés dans la panicule, seuls des épillets simples sont formés dans l'épi (Doebley, 1995b), ces épillets durcissent ensuite pour former une partie du fruit des graines de téosinte. L'épi de maïs semble régénérer l'état ancestral, la conséquence est que le maïs forme davantage de fleurs femelles et donc de graines.
- L'épi de maïs présente au minimum 4 rangées de grains alors que celui de téosinte n'en a que 2.
- Le rachis (l'axe de l'épi) du maïs reste intact alors qu'il se désarticule chez la téosinte (en raison de la présence de couches d'abscission), permettant la dispersion des graines. Pour l'Homme la récolte est facilitée et le mais devient dépendant de l'Homme pour se reproduire.
- Les glumes (la feuille à la base d'une fleur) du maïs sont petites et molles, tandis que celles de la téosinte (plus la cupule, c'est à dire le fragment associé du rachis) entourent complétement chaque graine. Les grains (fruits cupulés) de la téosine résistent au passage dans le dans le tube digestif des Oiseaux et des Mammifères qui les dispersent (Wilkes, 1967). Au contraire, les grains du maïs sont exposés aux prédateurs. Il existe une relation entre ce caractère et le précédent: la domestication entraine un changement dans le développement de l'épi de sorte que les cupules et les glumes forment son axe interne au lieu d'envelopper les graines, une sorte de retournement (Wang, 2005).
- La floraison du maïs est limitée dans le temps et les fleurs femelles apparaissent après les fleurs mâles, ce qui favorise la fécondation croisée.
L'augmentation de la taille des fruits ou des graines, la réduction de la dispersion des semences et la réduction de la ramification sont des caractéristiques communes aux plantes domestiquées (Stitzer, 2018).
On considère aujourd'hui que parviglumis est la variété ancestrale, mais des échanges semblent avoir eu lieu ultérieurement avec mexicana, plus adaptée à l'altitude, puisque vivant sur les haut-plateaux mexicains (Yang, 2023). La domestication a du prendre beaucoup de temps d'autant que se nourrir essentiellement de Maïs représente un régime très déséquilibré du point de vue des acides aminés essentiels (le Maïs manque de lysine et de tryptophane par rapport aux besoins humains) qui n'est rétabli qu'avec la consommation simultanée du Haricot ou de la Courge. Les paléoanthropologues pensent que le maïs a probablement été d'abord utilisé pour produire une boisson fermentée (chicha) plutôt que comme nourriture (Sallmann, 2022 p.35) d'autant que la tige de la téosinte est sucrée.
En 2018, Logan Kistler et ses collaborateurs proposent une évolution des premières variétés semi-domestiquées de Maïs (crées au Mexique dans la vallée tropicale humide de Balsas, vers 9 000 BP) sur les territoires du Pérou et de la Bolivie (en isolation des souches ancestrales de téosinte du Mexique) vers 6 000 BP, avant leur retour en Amérique centrale lors de vagues de migrations successives (Kistler, 2018).
Les études (analyses isotopiques de la dentition) menées dans un abri sous-roche du Belize par Douglas Kennett et ses collaborateurs montrent que le maïs n'a pris une part significative (70%) dans l'alimentation qu'il y a 6 000 ans. Entre 6 700 BP et 6 000 BP il apparait constituer une part minoritaire (autour de 30%) de l'alimentation et seulement chez certains individus (Kennett, 2020); il n'est pas du tout consommé avant 6 700 BP. Comme le patrimoine génétique des Mayas provient au moins à 50% de migrants d'Amérique du sud, on estime que ces migrants ont apporté avec eux des variétés de Maïs davantage domestiquées que celles préalablement présentes en Amérique centrale.
Le Maïs se répand dans l'ouest de l'Amérique vers 4 000 BP, en Louisiane vers 3 000 BP (Sallmann, 2022 p.35) et dans les Caraïbes où il est découvert par Christophe Colomb en 1493. La diffusion vers le nord s'accompagne de changements significatifs dans les QTL contrôlant la photopériode.
En Europe, le maïs devient généralement un complément alimentaire mais plus souvent il sera un aliment pour le bétail. Quelques régions l’adoptent comme aliment de base, avec des préparations spécifiques comme la polenta en Italie ou la mamaliga en Roumanie.
↑ Ning Yang et al. . 2023. Two teosintes made modern maize. Science 382, eadg8940. DOI: 10.1126/science.adg8940
↑ Jean-Michel Sallmann. 2022. L'Amérique du nord - 25 000 ans av. notre ère - 19e siècle. Belin.
Les amérindiens d'Amérique du nord.
↑ Douglas J. Kennett et al.. 2020. Early isotopic evidence for Maize as a staple grain in the Americas. Sci. Adv.6, eaba3245. DOI: 10.1126/sciadv.aba3245.
↑ Logan Kistler et al.. 2018. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 362: 1309-1313. DOI: 10.1126/science.aav0207.
↑ Michelle C. Stitzer, Jeffrey Ross-Ibarra. 2018. Maize domestication and gene interaction. New Phytologist. DOI: 10.1111/nph.15350
Un article essentiel avec une excellente synthèse historique.
↑ John Harshberger. 1896. Fertile crosses of teosinte and maize. Garden and Forest, 9 (1907): pp.398-402. archive.org
↑ Charles Darwin. 1868. Animals and Plants under Domestication. Orange Judd & Co., New York, USA, vol.1 pp.385-387 (archive.org).
↑ Jean-Michel Sallmann. 2022. L'Amérique du nord - 25 000 ans av. notre ère - 19e siècle. Belin.
Les amérindiens d'Amérique du nord.
↑ Douglas J. Kennett et al.. 2020. Early isotopic evidence for Maize as a staple grain in the Americas. Sci. Adv.6, eaba3245. DOI: 10.1126/sciadv.aba3245.
↑ Logan Kistler et al.. 2018. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 362: 1309-1313. DOI: 10.1126/science.aav0207.
↑ Michelle C. Stitzer, Jeffrey Ross-Ibarra. 2018. Maize domestication and gene interaction. New Phytologist. DOI: 10.1111/nph.15350
Un article essentiel avec une excellente synthèse historique.
↑ John Harshberger. 1896. Fertile crosses of teosinte and maize. Garden and Forest, 9 (1907): pp.398-402. archive.org
↑ Charles Darwin. 1868. Animals and Plants under Domestication. Orange Judd & Co., New York, USA, vol.1 pp.385-387 (archive.org).
La domestication du Lin ↑
Le Lin cultivé, Linum usitatissimum, est étroitement apparenté au Lin sauvage, Linum bienne avec lequel il est interfertile. Le classement en deux espèces différentes peut donc être discuté, même si le fait que les graines du lin cultivé dépendent de l'Homme pour leur dispersion, le fruit en capsule ne s'ouvrant pas à maturité (une des caractéristiques de la domestication). C'est une plante autogamme (autopollinisation très majoritaire).Le Lin est une des premières plantes domestiquées. L'histoire est complexe car le lin a deux utilisations: pour son huile et pour ses fibres. Les études génétiques, encores assez limitées semblent indiquer une domestication unique, dans le croissant fertile (Syrie), vers 12 000 à 11 000 BP (Hillman, 1975). La teneur en huile augmente la première, puis la taille des graines vers 8 000 BP. Les variétés textiles présentent des plants plus grands (environ 1m) et moins ramifiés avec des graines plus petites.
Une plante à tout faire
Le Lin était une plante très importante en Egypte, à la fois pour son usage alimentaire et la fabrication de vêtements. La tapisserie de Bayeux datant du 11e siècle est en lin et la plante était en plus considérée en Europe comme une plante magique. La culture a régressé en France avec l'expulsion des Huguenots (1685) experts dans sa culture (qui demande beaucoup de savoir-faire). Au 18e siècle, le coton a considérablement remplacé le lin dans les usages textiles en Europe.
Pour l'usage alimentaire, si la graine se conserve bien, l'huile se dégrade rapidement et doit être préservée de la lumière et de l'oxygène; de ce fait elle a été interdite en France pour l'alimentation humaine de 1908 à 2010; les graines entières résistent à la digestion, mais les graines broyées ou moulues sont aujourd'hui présentes dans des baguettes de pain "haut de gamme". On redécouvre aujourd'hui un rôle médicinal aux graines broyées, pour leur richesse en acides gras oméga-3, mais peut-être aussi pour d'autres composants.
L'huile a toujours été utilisée dans l'industrie (peintures, protection du bois, lubrifiant). Et on redécouvre qu'avec ses fibres extrèmement résistantes et fortement isolantes, le lin est une plante à tout faire. Les fibres ont été utilisées pour fabriquer des cordages, se prêtent à la réalisation de papiers fins, d'isolants; les déchets peuvent remplacer le bois pour l'usage calorifique.
Des origines du Lin cultivé.
G. Hillman. 1975. The plant remains from Tell Abu Hureyra: A preliminary report. Proc Prehist Soc 41:70–73.
G. Hillman. 1975. The plant remains from Tell Abu Hureyra: A preliminary report. Proc Prehist Soc 41:70–73.
La domestication du Riz ↑
Les indices de domestication du Riz sont moins spectaculaires que ceux présentés par le Maïs; on retiendra surtout le maintien des grains sur la plante (Fuller, 2009) qui relève du gène sh4 (Li 2006), mais aussi le gène OsSHATTERING1 (OsSh1), exprimé à la base de la fleur et encodant un facteur de transcription à homeo domaine impliqué dans la formation d'une couche d'abscission (Konishi, 2006). Le changement permet la récolte et rend le Riz dépendant de l'Homme pour sa reproduction.Le riz cultivé en Asie et les espèces sauvages proches possèdent 12 chromosomes (2n=24). Le génome est de petite taille comparativement à d'autres graminées et c'est une des raisons qui a permis le séquençage complet du génome en 2005, une première pour les plantes domestiquées. Le riz cultivé originellement en Asie (et depuis dans le monde entier) est classé dans l'espèce Oryza sativa avec deux groupes principaux Oryza sativa ssp indica et Oryza sativa ssp japonica. Les deux variétés différent par de nombreux caractères mais la distinction la plus flagrante est culinaire, indica étant moins collant que japonica (ce dernier est plus facile à consommer avec des baguettes).

Oryza rufipogon, Oryza nivara et Oryza sativa ssp japonica (Nipponbare)
d'après Chun-Yan Jing, 2023, fig.1 (Nature).
d'après Chun-Yan Jing, 2023, fig.1 (Nature).
O.sativa possède une diversité génétique considérable (nombreuses sous-espèces: International Rice Genebank possède plus de 100 000 variétés) et est proche des variétés sauvages classées dans les espèces O.rufipogon (grande, pérenne et inféodée aux milieux humides) et O.nivara (petite, annuelle et plus adaptée aux milieux secs). L'autofécondation domine chez O.nivara; les deux modes de fécondation sont également fréquents chez O.rufipogon; toutes ces espèces sont susceptibles de s'hybrider (ce qui fait que le classement actuel en espèces séparées pourrait être remis en cause).

Les restes de riz fossilisés dans la vallée du Yangtsé sont datés entre 13 900 BP et 8 000 BP; les flèches noires représentent la diffusion des variétés domestiquées, argumenté par l'archéologie et la linguistique. Une autre région de domestication potentielle est la vallée du Gange avec des datations de riz fossiles moins anciennes (d'après Michael Kovaxh, 2007, repris de Peter Bellwood).
L'histoire de la domestication du riz asiatique est apprement discutée
Deux thèses s'affontent: celle d'une domestication unique en Chine et celle de domestications séparées en Chine et en Inde; les archéologues chinois et indiens se disputant depuis près d'un siècle la parternité de la domestication.Selon les études génétiques, de nombreuses caractéristiques de indica et japonica divergent antérieurement aux données archéologiques sur la domestication, ce qui crée un paradoxe.
Yingqing Lu et al. (Lu, 2022) proposent une hybridation naturelle entre O.rufipogon et O.nivara suivie d'une première phase de domestication. Une deuxième phase de domestication beaucoup plus diversifiée géographiquement conduit à une relative sépartion des variétés indica et japonica à partir d'un pool de variations préexistantes dans l'espèce domestiquée initialement. Et en 2023 Chun-Yan Jing (Jing, 2023) reprend la proposition de Peter Civáň (Civáň, 2015) impliquant 3 centres de domestication:

Centres de domestication hypothétiques du riz asiatique d'après Jing, 2023. Les losanges verts indiquent les allèles domestiqués dans japonica (~80%) et les triangles bruns ceux domestiqués dans indica (~20%). L'événement de domestication positionné en Asie du sud-est demande des études ultérieures (Crédit IBCAS).
La culture du riz, surtout celle de la variété Oryza sativa ssp japonica, en rizière inondée (et surtout cultivée sur de petites surfaces, peu mécanisées), demande un travail considérable.
O.rufipogon est considéré comme une espèce invasive dans les champs de riz aux Etats-Unis; comme il perd l'essentiel de ses grains sur le sol avant la récolte, ce n'est donc pas la présence de ses grains dans le riz récolté qui est gênante, mais la perte de rendement.
↑ Chun-Yan Jing et al. 2023. Multiple domestications of Asian rice. Nat. Plants 9: 1221–1235. DOI: 10.1038/s41477-023-01476-z, figure 1, Supplementary information.
↑ Yingqing Lu et al.. 2022. Early Domestication History of Asian Rice Revealed by Mutations and Genome-Wide Analysis of Gene Genealogies. Rice 15, 11. DOI: 10.1186/s12284-022-00556-6.
↑ Michela Osnato et al.. 2022. Evolution of flowering time genes in rice: from the paleolithic to the anthropocene. Plant Cell Environment, volume46, issue4: 1046-1059. DOI: 10.1111/pce.14495.
↑ Ran Xu, Chuanqing Sun. 2021. What happened during domestication of wild to cultivated rice. The Crop Journal, Vol.9, Issue 3: 564-576. DOI: 10.1016/j.cj.2021.02.005
↑ Ran Xu, Chuanqing Sun. 2021. What happened during domestication of wild to cultivated rice. The Crop Journal, Vol.9, Issue 3: 564-576. DOI: 10.1016/j.cj.2021.02.005
↑ André Gallais. 2016. L'hybridation dans le monde végétal.
↑ P. Civáň et al. 2015. Three geographically separate domestications of Asian rice. Nature Plants 1, 15164. DOI: 10.1038/nplants.2015.164.
↑ Ewen Callaway. 2014. Domestication: the birth of rice. Nature 514, S58–S59. DOI: 10.1038/514S58a.
↑ Jeanmaire Molina et al.. 2011. Molecular evidence for a single evolutionary origin of domesticated rice. PNAS 108 (20): 8351-8356. DOI: 10.1073/pnas.110468610.
↑ Dorian Q. Fuller et al.. 2009. The domestication process and domestication rate in rice: spikelet bases from the lower Yangtze. Science 323: 1607–1610. DOI: 10.1126/science.1166605.
↑ Duncan Vaughan et Norihiko Tomooka. 2008. The Evolving Story of Rice Evolution. Plant Science 174: 394-408. DOI: 10.1016/j.plantsci.2008.01.016.
↑ Changbao Li et al.. 2006. Rice domestication by reducing shattering. Science 311: 1936-1939. DOI: 10.1126/science.1123604.
Brigitte Courtois. Une brève histoire de l'amélioration génétique du riz.
↑ Yingqing Lu et al.. 2022. Early Domestication History of Asian Rice Revealed by Mutations and Genome-Wide Analysis of Gene Genealogies. Rice 15, 11. DOI: 10.1186/s12284-022-00556-6.
↑ Michela Osnato et al.. 2022. Evolution of flowering time genes in rice: from the paleolithic to the anthropocene. Plant Cell Environment, volume46, issue4: 1046-1059. DOI: 10.1111/pce.14495.
↑ Ran Xu, Chuanqing Sun. 2021. What happened during domestication of wild to cultivated rice. The Crop Journal, Vol.9, Issue 3: 564-576. DOI: 10.1016/j.cj.2021.02.005
↑ Ran Xu, Chuanqing Sun. 2021. What happened during domestication of wild to cultivated rice. The Crop Journal, Vol.9, Issue 3: 564-576. DOI: 10.1016/j.cj.2021.02.005
↑ André Gallais. 2016. L'hybridation dans le monde végétal.
↑ P. Civáň et al. 2015. Three geographically separate domestications of Asian rice. Nature Plants 1, 15164. DOI: 10.1038/nplants.2015.164.
↑ Ewen Callaway. 2014. Domestication: the birth of rice. Nature 514, S58–S59. DOI: 10.1038/514S58a.
↑ Jeanmaire Molina et al.. 2011. Molecular evidence for a single evolutionary origin of domesticated rice. PNAS 108 (20): 8351-8356. DOI: 10.1073/pnas.110468610.
↑ Dorian Q. Fuller et al.. 2009. The domestication process and domestication rate in rice: spikelet bases from the lower Yangtze. Science 323: 1607–1610. DOI: 10.1126/science.1166605.
↑ Duncan Vaughan et Norihiko Tomooka. 2008. The Evolving Story of Rice Evolution. Plant Science 174: 394-408. DOI: 10.1016/j.plantsci.2008.01.016.
↑ Changbao Li et al.. 2006. Rice domestication by reducing shattering. Science 311: 1936-1939. DOI: 10.1126/science.1123604.
Brigitte Courtois. Une brève histoire de l'amélioration génétique du riz.
La domestication séparée du riz africain fait consensus
Il existe un autre riz cultivé en Afrique, Oryza glaberrima qui s'hybride difficilement avec sativa et dérive de l'espèce sauvage O. barthiiNat Genet. 2014; 46(9): 982–988. Published online 2014 Sep 1. doi: 10.1038/ng.3044 PMCID: PMC7036042 PMID: 25064006 The genome sequence of African rice (Oryza glaberrima) and evidence for independent domestication Muhua Wang,#1
Philippe Cubry et al. The rise and fall of African rice cultivation revealed by analysis of 246 new genomes, Current Biology , 5 juillet 2018.
La domestication du Sorgho ↑
La systématique des sorghos a longtemps été complexe, à cause d’une énorme variabilité des formes à l’échelle du continent, conduisant à la description de dizaines d’espèces dans le genre Sorghum. 5 races avaient été reconnues, bicolor, kafir, caudatum, durra et guinea, qui finalement ont été réunies, avec toutes les espèces cultivées « à grains », sous le binôme S. bicolor, ssp. bicolorLa domestication du Soja ↑
Le Soja est utilisé en Asie depuis 5000ans; il est longtemps resté inconnu ailleurs. La production a considérablement augmenté à la fin du 20e siècle en particulier en Amérique.Le Soja est une plante largement autogamme. La diversité génétique est grande entre ses multiples lignées (variétés), un peu plus faible à l'intérieur de chaque lignée. de fait la vigueur hybride s'applique assez peu au Soja.
La domestication de la Pomme de Terre ↑
Histoire de la pomme de terre. Wikipedia.
Solanum berthaultii. Wikipedia.
Plante endémique de la région andine de la Bolivie
Solanum berthaultii. Wikipedia.
Plante endémique de la région andine de la Bolivie
Le sucre: histoire d'une addiction: canne à sucre et Betterave (détails) ↑
«La canne à sucre moderne est, parmi les plantes cultivées, celle qui a le génome le plus complexe, car elle a un nombre de chromosomes très élevé qui proviennent de plusieurs espèces. Cette complexité a ralenti la compréhension de son origine, de sa diversité et de l’architecture de son génome.»Angélique D’Hont Coordinatrice de l’étude, Cirad - UMR Agap
on estime aujourd'hui que l'ancêtre sauvage ayant joué le rôle majeur est Saccharum robustum présent en Nouvelle Guinée, mais qui contient très peu de sucre. La variété domestiquée est transportée vers l'Inde, la Chine et vers des iles de Mélanésie. La domestication a augmenté considérablement la présence de sucre et a adapté la plante à des climats moins chauds. Elle est facilement propagée par boutures.
Garsmeur et al.. 2025. The genomic footprints of wild Saccharum species trace domestication, diversification, and modern breeding of sugarcane. Cell , 10.1016/j.cell.2025.09.017. ANSES.
Betterave sucrière. Wikipedia.
Solanum berthaultii. Wikipedia.
Plante herbacée du bord de mer
Betterave sucrière. Wikipedia.
Solanum berthaultii. Wikipedia.
Plante herbacée du bord de mer
La domestication du Coton (détails) ↑
Une des rares plantes qui n'est pas domestiquée en tant qu'aliment. C'est une histoire complexe, car il n'y a pas un, mais des cotons. Et donc plusieurs domestications (4).Gossypium arboreum est une espèce domestiquée dans la vallée de l'Indus au Pakistan et en Inde; le site de Mehrgarh a livré des fibres et des graines datant de 8000 BP. G. arboreum est est introdute au Moyen orient (250 BP) et en Afrique du nord (1500 BP). On pense que G. arboreum a été introduit en Chine comme plante ornementale vers 1000 BP. G. arboreum est encore couramment cultivée dans les petites iles d'Indonésie.
Gossypium herbaceum, originaire d'Afrique (ou de l'Indus ?), a été cultivée en Arabie et en Syrie.
Gossypium hirsutum représente 90 % de la production mondiale; son utilisation quasi exclusive date du 19e siècle. G. hirsutum était à l'origine cultivé au Mexique (Yucatán, 3400 BP) mais pourrait être issu de G. barbadense (coton "égyptien" ou Pima) présent au Pérou où des traces archéologiques datant de 4500 BP ont été découvertes à Caral. G. barbadense est un coton de haute qualité à longues fibres son nom de coton égyptien vient de l'histoire de sa culture au 19e siècle et il est toujours cultivé en Egypte.
G. arboreum et G. herbaceum s'hybrident et donnent des descendants fertiles mais les graines germent mal et les plantes hybrides retournnent rapidement aux phénotypes parentaux (Christopher Viot, 2019, Domestication and varietal diversification of Old World cultivated cottons).
L'ancêtre commun de G. arboreum et G. herbaceum est estimé à 0,4-2,5 million d'années; la fourchette est large mais la séparation est de toute façon bien antérieure à la domestication.
La domestication a permis de transformer les cotons sauvages, sensibles à la photopériode, en plantes annuelles domestiques, moins réactives à la photopériode. Les cotons anciens pouvaient être bien moins exigeants en eau et la culture comme plante annuelle contournait la sensibilité au froid. En dehors des fruits récoltés à la main à l'origine, les feuilles et les racines pouvaient servir de combustible ou de nourriture pour le bétail avec la réserve qui suit: les graines sont riches en huile, mais contiennent le gossypol, toxique pour les humains et les non-ruminants (Charlène Bouchaud, 2019, Tightening the thread from seed to cloth). De nos jours il est possible d'éliminer le gossypol par un traitement chimique.
Les premiers cotons cultivés sont résistants à la sécheresse et aux ravageurs (Kulkarni et al. 2009); les variétées cultivées de G. arboreum sont généralement bien adaptées aux climats de mousson (Valicek 1979); G. herbaceum var. acerifolium supporte des climats plus froids et plus secs (Hutchinson et al. 1947). De nombreuses variétés de G. arboreum étaient cultivées en Inde, adaptées aux climats locaux (Hutchinson et al. 1947, Kulkarni et al. 2009). Les problèmes rencontrées par les hybrides et les hybrides OGM commercialisés issus de G. hirsutum par les semenciers ne semblent pas présenter ces qualités. La fécondation est généralement une autofécondation, mais le pollen (assez lourd) peut-être transporté par les insectes.
Le coton doit être filé (fuseau et son fusaïole) et les tissus de coton se décomposent en 3 semaines dans un sol humide. (Christopher Viot, 2019, Domestication and varietal diversification of Old World cultivated cottons).
La fibre prend bien la teinture et a permis le développement de toutes sortes de traditions textiles.
L'histoire de la culture du coton, comme celle de la production du sucre est une histoire globale sans doute plus économique et politique que biologique. Elle s'entrecroise avec celle de l'industrie textile et de la colonisation et représente (avec l'histoire du sucre) un des premiers exemples développé de mondialisation.
La production mondiale de coton a considérablement augmenté ces dernières années et l'Inde est devenu le premier producteur. Le coton biologique (cultivé sans pesticides et donc non OGM, référentiel GOTS (Global Organic Textile Standard).) ne représente que 0,2% de la production mondiale totale (83 millions de tonnes en 2020) et dans la fabrication du tissu (suivant le label) il est possible d'accepter un certain pourcentage de fibres non biologiques.
L'histoire de la domestication du coton (Gossypium) . EGFR.
Charlène Bouchaud et Elsa Yvanez (dir.). 2019. Coton de l’Ancien Monde: domestication, culture, usages et échanges. Revue d’Ethnoécologie. En ligne. MNHN. DOI: 10.4000/ethnoecologie.4063
Christopher Viot Domestication and varietal diversification of Old World cultivated cottons (Gossypium sp.) in the Antiquity in Charlène Bouchaud et Elsa Yvanez (dir.). 2019.
Coton. Wikipedia.
Histoire de la culture du Coton. Wikipedia.
Charlène Bouchaud et Elsa Yvanez (dir.). 2019. Coton de l’Ancien Monde: domestication, culture, usages et échanges. Revue d’Ethnoécologie. En ligne. MNHN. DOI: 10.4000/ethnoecologie.4063
Christopher Viot Domestication and varietal diversification of Old World cultivated cottons (Gossypium sp.) in the Antiquity in Charlène Bouchaud et Elsa Yvanez (dir.). 2019.
Coton. Wikipedia.
Histoire de la culture du Coton. Wikipedia.
La guerre des semences ↑
Le Brésil ou en Argentine par exemple ne reconnaissent pas de droits de propriété pour les semences.GURT (Genetic Use Restriction Technology), Terminator et Traitor. Il s'agit de modifier les semences pour les rendre stériles. En 2000, la Convention des Nations unies sur la diversité biologique (CDB) a adopté un moratoire de fait sur ces technologies, ce qui n'empêche pas un pays de l'utiliser, mais empêche de l'imposer à ses voisins. Ce moratoire reste fragile.
Plantes OGM ↑
(en cours de rédaction).Et vous n'avez encore rien vu
Les OGM de seconde génération constituent un autre sujet dont les perspectives sont tellement diverses et imprévisibles qu'elle sortent du cadre de cette page; consultez plutôt: Brave New World.
Les organismes génétiquement modifiés (OGM). Ministère de l'agriculture.
Une synthèse intéressante en particulier comme mise au point sur la réglementation.
List with GMOs and their identifiers. Euginius (European GMO Initiative for a Unified Database System).
Une synthèse intéressante en particulier comme mise au point sur la réglementation.
List with GMOs and their identifiers. Euginius (European GMO Initiative for a Unified Database System).